












- DAZ.online
- DAZ / AZ
- DAZ 8/2011
- Immunbiologische Effekte ...
Mikronährstoffe
Immunbiologische Effekte von Mikronährstoffen
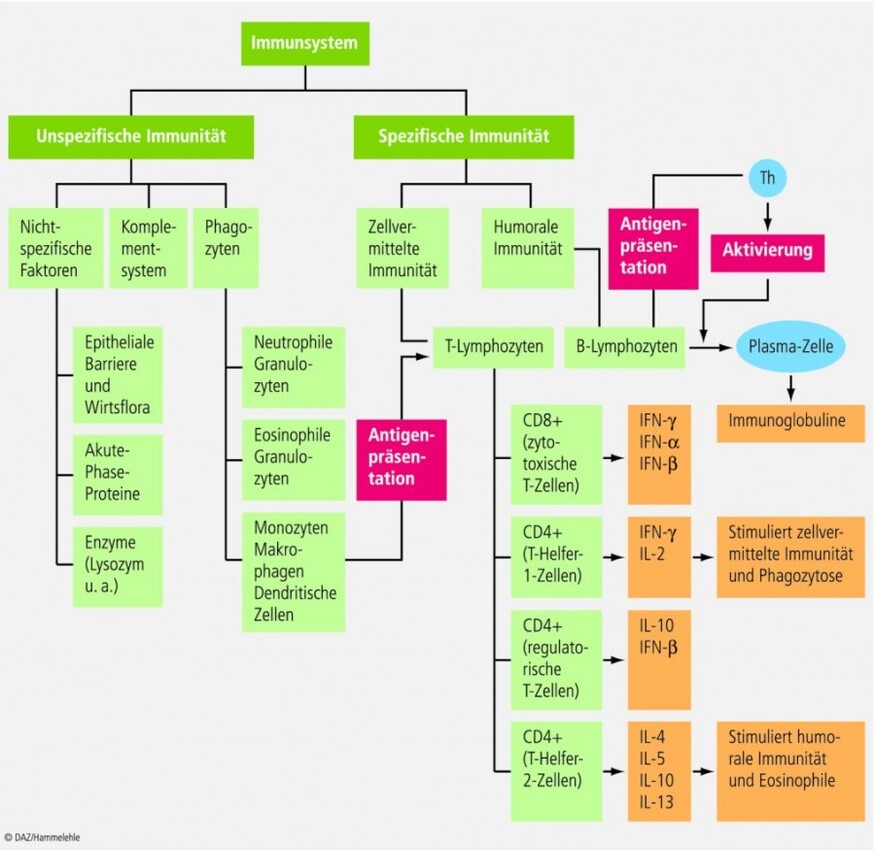
Das Immunsystem ist ein diffus strukturiertes und über den gesamten Organismus verteiltes Organnetzwerk, das aus zellulären und humoralen Faktoren besteht. Diese vermitteln sowohl die antigen-unspezifische (angeborene) als auch die antigen-spezifische (erworbene) Immunität des Organismus (Abb. 1). In Abhängigkeit von seiner Aktivierbarkeit wird zwischen der unspezifischen und der spezifischen Immunität differenziert:
Unspezifisches Immunsystem: Hierbei handelt es sich um ein phylogenetisch hochkonserviertes Abwehrsystem, das zur unmittelbaren Immunantwort befähigt und für die erste Phase einer Immunreaktion verantwortlich ist. Es wird direkt, d. h. ohne Vorprozessierung von Antigenen aktiviert und ist – worauf die Bezeichnung bereits hindeutet – in seiner Wirkung antigen-unspezifisch. Neben den zellulären Komponenten (Tab. 1) besteht das unspezifische Immunsystem aus humoralen Faktoren; hierzu zählen u. a. Effektorproteine und lokal wirksame Mediatoren wie Zytokine und Akut-Phase-Proteine (Tab. 2).
Tab. 1: Überblick zur Funktion und Wirkungsweise der an der unspezifischen Immunabwehr beteiligten zellulären Faktoren. | ||
Immunzelltyp |
Aktivierung |
Effekt |
Neutrophile Granulozyten (syn.: polymorphonukleäre Granulozyten) |
|
|
Monozyten/Makrophagen und dendritische Zellen |
|
|
Eosinophile Granulozyten |
|
|
Basophile Granunlozyten |
|
|
Mastzellen |
|
|
Tab. 2: Überblick zur Funktion und Wirkungsweise der an der unspezifischen Immunabwehr beteiligten humoralen Faktoren. | ||
Element |
Bildungsort |
Effekt |
Zytokine | ||
|
|
|
|
|
|
Akut-Phase-Proteine | ||
|
|
|
|
|
|
"Alternatives" Komplementsystem | ||
|
|
|
|
|
|
|
|
|
Spezifisches Immunsystem: Dieses stellt das phylogenetisch jüngere System der Immunabwehr dar, ist selektiv und damit antigenspezifisch wirksam sowie für die zweite Phase der Immunreaktion verantwortlich. Es wirkt indirekt, d. h. Voraussetzung für seine Aktivität ist die Vorprozessierung von Antigenen durch das unspezifische System. Funktional lassen sich auch hier zelluläre und humorale Komponenten unterscheiden:
-
Das zelluläre System basiert auf B- und T-Lymphozyten, die beim Erwachsenen zusammen eine Masse von etwa 1 kg ausmachen. Sowohl B- als auch T-Lymphozyten entstammen dem hämopoetischen Gewebe. Erstere differenzieren im Thymusgewebe, Letztere in der Leber (vor der Geburt) bzw. im Knochenmark (nach der Geburt) zu antigenreaktiven Immunzellen. 75% der mononukleären Zellen im Blut zählen zu den T-Lymphozyten; 25% zu den B-Lymphozyten. Strukturell lassen sich die beiden Lymphozytenpopulationen durch ihre spezifischen Oberflächenmarker differenzieren. T-Lymphozyten sind durch die Expression des Oberflächenrezeptors CD3, B-Lymphozyten durch die Oberflächenrezeptoren CD19 und CD20 charakterisiert (Tab. 3).
Tab. 3: Überblick zur Funktion und Wirkungsweise der an der spezifischen Immunabwehr beteiligten
zellulären Faktoren.ImmunzelltypAktivierungEffektT-Lymphozyten- Naive T-Zellen
- IL-12 und IL-4
- Vorstufe von T-Effektorzellen (T-Helfer oder zytotoxische T-Zelle)
- T-Helferzelle (Th1- und Th2Zellen; CD4+)
- Über Bindung eines von antigen-prozessierenden Zellen (z. B. Makrophagen) präsentierten Antigens an den T-Zellrezeptor
- Verstärkung der B-Zellfunktion und Produktion spezifischer Antikörper (= Induktion der humoralen Immunantwort)
- Makrophagenaktivierung
- Zytotoxische T-Zellen (CD8+)
- Über Bindung des T-Zellrezeptors an das Zielantigen
- Lysierung von Zellen (Tumor-, Bakterien- und virusinfizierte Zellen), die ein an den MHC-I-Komplex gebundenes Antigen aufweisen.
- Induktion der Apoptose in Zielzellen
- Regulatorische T-Zellen (CD4+)
- Dämpfung von Immunreaktionen
- T-Gedächtniszellen (CD4+)
- Rasche Immunantwort bei erneutem Antigenkontakt
B-Lymphozyten- B-Zellen
- T-Zell-abhängig (Proteinantigene): über T-Helferzellen
- T-Zell-unabhängig (Polysaccharid-antigene): über direkten Antigenkontakt
- Bindung von Antigenen über Antigenrezeptor
- Antigenpräsentation
- B-Plasmazellen
- T-Helferzelle
- Immunglobulinsynthese
- Antikörperabhängige zellvermittelte Zytotoxizität
- B-Gedächtniszellen
- T-Helferzelle
- Rasche Immunantwort bei erneutem Antigenkontakt
-
Das humorale System besteht aus Zytokinen und Immunglobulinen (Antikörpern). Die humorale Achse der spezifischen Immunität wird über B-Lymphozyten vermittelt, wobei die humorale Abwehrreaktion im Fall der meisten Antigene die synergistische Wirkung von T-Zellen erforderlich macht. Nähere Details zur Funktion wichtiger humoraler Mediatoren, die an der spezifischen Immunität beteiligt sind, siehe Tab. 4.
Tab. 4: Überblick zur Funktion und Wirkungsweise der an der spezifischen Immunabwehr beteiligten
humoralen Faktoren.ElementBildungsortEffektZytokine- Interleukin-2 (IL-2)
- Aktivierte T-Zellen
- T-Zellproliferation und ‑Differenzierung↑
- Aktivierung von zytotoxischen T-Zellen und von Makrophagen
- Interleukin-4 (IL-4)
- CD4+-T-Zellen
- Mastzellen
- B-Zellaktivierung
- Ig-Klassenwechsel von IgM auf IgG ung IgE ("Isotopen-switch")
- Interleukin-5 (IL-5)
- CD4+-T-Zellen
- Mastzellen
- Proliferation und Differenzierung von Eosinophilen
- Proliferation und Differenzierung aktivierter T- und B-Zellen
- Interferon γ (IFNγ)
- CD4+-Zellen
- NK-Zellen
- Expression von MHC-Klasse-II-Molekülen
- Makrophagenaktivierung
- Adhäsion von Leukozyten an das Endothel
Immunglobuline- IgE
- Plasmazellen
- Schutz vor Darmparasiten
- Antikörperabhängige zelluläre Zytotoxizität
- Vermittelt allergische Reaktionen
- IgG
- Plasmazellen
- Virus- und Toxinneutralisation
Wie bei allen Körperorganen ist auch die Funktionsweise des Immunsystems an eine adäquate Energie- und Nährstoffsubstratzufuhr gekoppelt. Dies umso mehr, da die einzelnen Komponenten des Immunsystems einem hohen Turnover unterliegen, so dass ihr Substratbedarf im Vergleich zu den meisten anderen Körpersystemen gesteigert ist. Nur bei einer bedarfsgerechten Zufuhr aller essentiellen Nährstoffe laufen die biochemischen und zellulären Mechanismen, die die Grundlage des Immunsystems bilden, optimal ab. Es kann daher nicht verwundern, dass die Immunkompetenz des Organismus entscheidend von der Zufuhr an Mikronährstoffen (Vitamine, Mineralstoffe und bestimmte Fettsäuren), beeinflusst wird (Tab. 5) [2 – 5]. Umgekehrt übt aber auch die Aktivität des Immunsystems Einflüsse auf den Nährstoffstatus aus, erkennbar u. a. bei immunologischen Erkrankungen wie dem aquired immunodeficiency syndrome (AIDS) [6 – 8] oder der rheumatoiden Arthritis [9 – 11]. Entsprechend gewinnen diätetische Maßnahmen auch bei Erkrankungen mit immunologisch-inflammatorischem Hintergrund an therapeutischer Relevanz [12 – 15].
Mikronährstoffe und Immunfunktion
Immunologisch aktive Zellen sind durch eine hohe Proliferationsrate charakterisiert, wodurch sie einen erhöhten Nährstoffbedarf aufweisen. Entsprechend laufen die biochemischen Prozesse, die die Grundlage des Immunsystems bilden, nur bei einer bedarfsgerechten Zufuhr aller Mikronährstoffe optimal ab. In diesem Zusammenhang fungieren Vitamine und Spurenelemente primär als [3, 5]:
Cofaktoren von Enzymen, die in die Replikation, Transkription und Translation sowie in die Signaltransduktion und damit in die Teilung und Proliferation der Immunzellen eingeschaltet sind (z. B. B-Vitamine, Zink);
Elemente des antioxidativen Systems, die für die Integrität von Immunzellen, die einer hohen Belastung an reaktiven Sauerstoffspezies (ROS) ausgesetzt sind, notwendig sind (Vitamine E, C, Selen);
Bestandteile von Transkriptionsfaktoren (z. B. Vitamine A, D), die als Kodeterminanten auf die Genexpression immunbiologisch relevanter Proteine Einfluss nehmen;
Modulatoren des epigenetischen Systems, die in die DNA-Methylierung (z. B. Folsäure) sowie die posttranslationale Modifikation (Biotinylierung via Biotin und Poly(ADP-ribosylation) via Niacin) von Histonen eingebunden sind.
Der aus immunbiologischer Sicht traditionell am besten bekannte Mikronährstoff ist das Vitamin C. Seit dem Erscheinen des von Linus Pauling (1901 – 1994) stammenden locus classicus "Vitamin C and the Common Cold" im Jahre 1970 [16], reißt die Diskussion um die Bedeutung dieses Vitamins für die Immunfunktion nicht ab. Insbesondere die Beantwortung der Frage, inwieweit Vitamin C für die Vorbeugung und Behandlung von Erkältungskrankheiten von Relevanz ist, fällt nach wie vor kontrovers aus [1].
Vitamin C – Struktur und Biochemie
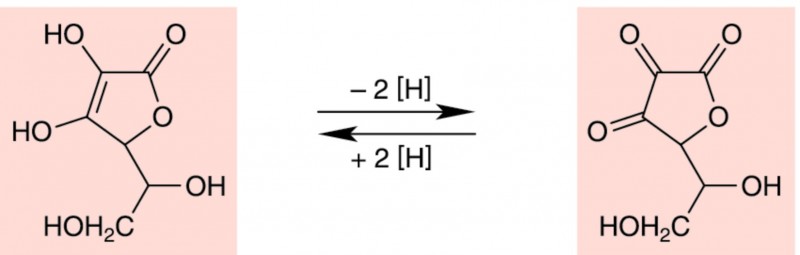
Vitamin C ist der Oberbegriff für L-Threo-hex-2-enono-1,4-lacton und seine Derivate mit L-Ascorbinsäure-Wirkung. Chemisch gesehen ist Vitamin C ein Gulonsäurelacton, das sich von der Glucuronsäure ableitet. Von den vier möglichen Stereoisomeren ist das L-Enantiomer die physiologisch aktive Form. Die charakteristische Eigenschaft der Ascorbinsäure ist ihre Fähigkeit, verschiedene Substanzen zu reduzieren, wobei sie reversibel in eine oxidierte Form, die Dehydroascorbinsäure, übergeht (Abb. 2). Dehydroascorbinsäure kann im Organismus wieder enzymatisch zu Ascorbinsäure reduziert werden und besitzt daher die volle Vitaminwirksamkeit.
Ascorbinsäure ist aufgrund ihrer stark reduzierenden Eigenschaften als Redoxsystem an zahlreichen Stoffwechselprozessen beteiligt. Von Bedeutung ist die Mitwirkung der Ascorbinsäure an Hydroxylierungsreaktionen, die durch Mono- und Dioxygenasen (Polyl 4-, Prolyl 3- und Lysyl Hydroxylase; Trimethyllysin und γ-Butyrobetain Hydroxylase, Dopamin β-Hydroxylase, 4-Hydroxyphenylpyruvat Dioxygenase und HIF Prolyl 4-Hydroxlase) katalysiert, dazu zählen u. a. [17 – 22]:
- die cotranslationale Hydroxylierung von Prolin- und Lysinresten des Procollagens im Zuge der Kollagensynthese,
- die Hydroxylierung von Tryptophan bei der Serotoninbildung,
- die Synthese von Catecholaminen,
- die α-Amidierung im Zuge der Synthese zahlreicher Peptidhormone (u. a. Gastrin, Cholecystokinin, Calcitonin, Vasopressin und Oxytocin) sowie
- die Hydroxylierung von β-Hydroxy-ε-N-Trimethyllysin und γ-Butyrobetain im Rahmen der endogenen Carnitinsynthese.
Als weitere physiologische, aber enzymunspezifische Funktionen der Ascorbinsäure sind zu nennen:
- Beteiligung bei der Eisenübertragung vom Transportprotein Transferrin auf das Eisenspeicherprotein Ferritin.
- Detoxifikation zahlreicher Verbindungen in der Leber über die Stimulation der Cytochrom-P450-Synthese.
- Förderung der intestinalen Aufnahme von Eisen, indem schlecht absorbierbares Fe3+ durch Ascorbinsäure in das besser verfügbare Fe2+ überführt wird bzw. dadurch, dass Ascorbinsäure der Bildung unlöslicher Eisen-Tannin- und -Phytatkomplexen entgegenwirkt.
- Beteiligung an der antioxidativen Abwehr im hydrophilen Kompartiment der Körperzellen.
Auf die immunbiologische Relevanz einiger dieser Ascorbat-abhängigen Funktionen wird im Abschnitt "molekulare Mechanismen von Vitamin C für die Immunkompetenz" im Detail eingegangen.
Vitamin C – immunbiologische Bedeutung
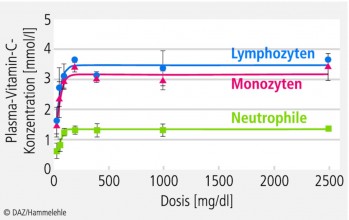
Bereits die Tatsache, dass immunkompetente Zellen (Lymphozyten, Neutrophile und Monozyten) mit 1 bis 4 mM eine Vitamin-C-Konzentration aufweisen, die etwa 10- bis 100-fach über den Vitamin-C-Plasmaspiegeln liegt, weist auf die immunologische Funktion von Vitamin C hin. Unterstrichen wird dies durch den Befund, wonach die o. g. Immunzellen den Mikronährstoff gegen einen Konzentrationsgradienten akkumulieren. Generell weisen mononukleäre Leukozyten etwa 2- bis 3-fach höhere Vitamin-C-Konzentrationen auf als polymorphnukleäre Zellen, wobei die Vitamin-C-Konzentration in Letzteren stärker mit den Serumwerten assoziiert sind, als dies bei mononukleären Leukozyten der Fall ist [3]. Zwischen der Vitamin-C-Zufuhr und der intrazellulären Ascorbat-Konzentration immunkompetenter Zellen wie Monozyten und Neutrophile besteht bei einmaliger oraler Gabe eine sigmoidale Abhängigkeit (Abb. 3). Eine vollständige intrazelluläre Sättigung wird bei Erwachsenen bei einer Vitamin-C-Zufuhr von etwa 100 mg/d beobachtet [23].
Die oben dargestellten Zusammenhänge unterstreichen die besondere Funktion, die Vitamin C für die zelluläre Immunität zukommt. Tatsächlich imponiert ein experimentell erzeugter Vitamin-C-Mangel durch Einschränkungen der zellulären, nicht aber der humoralen Immunabwehr [3]. So fand sich in einer Studie an gesunden Männern, deren Vitamin-C-Zufuhr über 60 Tage von 250 auf 5, 10 oder 20 mg pro Tag reduziert wurde, eine verminderte Immunantwort bei der Hautreaktion vom verzögerten Typ, einem Indikator der Immunfunktion. Die Immunantworten fielen auch nach dreiwöchiger Erhöhung auf 60 mg/d oder auf 250 mg/d noch niedriger aus als vor Versuchsbeginn [24].
Bedeutung von Vitamin C für die Aktivität von Phagozyten
Während Vitamin C keinen Einfluss auf die Anzahl der Phagozyten ausübt, wird die Aktivität dieser Immunzellen offenbar von der intra- und extrazellulären Vitamin-C-Konzentration beeinflusst. Folgende Befunde sind hierbei von Relevanz [1, 25, 26]:
Die Phagozytoseaktivität ist unter suboptimaler Vitamin-C-Zufuhr vermindert, wie die Mehrzahl der tierexperimentellen Studien zeigt, wenngleich auch gegenteilige Ergebnisse vorliegen.
Die Chemotaxis der Phagozyten wird von der Vitamin-C-Konzentration beeinflusst. Während die Ascorbat-induzierte Steigerung der Chemotaxis in vitro unphysiologisch hohe Vitaminmengen zu erfordern scheint, ist ein Effekt von Vitamin C auf die phagozytäre Chemotaxis unter in-vivo-Bedingungen indirekt möglich. Und zwar über eine Senkung des Histaminspiegels im Blut, wie sie nach Vitamin-C-Gabe beim Menschen zu beobachten ist.
Ein Abfall der intrazellulären Vitamin-C-Konzentration in Leukozyten ist mit einer Einschränkung ihrer Immunaktivität assoziiert.
Sowohl eine nicht-kontrollierte als auch zwei placebokontrollierte Interventionsstudien an Gesunden belegen, dass eine Vitamin-C-Supplementierung im Bereich von 1500 bis 3000 mg/d (Interventionszeitraum: Wochen bis Monate), die Lymphozytenproliferation zu steigern vermag, wobei dieser Effekt erst nach Adjustierung um Veränderungen der T-Zellzahl einen Trend zur statistischen Signifikanz (P = 0,07) erkennen ließ.
Zwischen der Vitamin-C-Konzentration des extrazellulären Mediums bzw. der Vitamin-C-Zufuhr und der Proliferation der Lymphozyten scheint eine dosisabhängige Beziehung zu bestehen: Eine physiologische Vitamin-C-Konzentration im Bereich von 5 bis 50 µg/ml verstärkt die Lymphozytenproliferation, während supraphysiologische Konzentrationen inhibierende Effekte zeigen.
Bei Erkrankungen, die mit einer Phagozytendysfunktion einhergehen (z. B. das Chediak-Higashi-Syndrom; siehe Kasten), lässt sich die Phagozytenaktivität durch Vitamin-C-Supplementierung verbessern.
Sowohl in-vitro- als auch tierexperimentelle Untersuchungen belegen die proliferationssteigernden Effekte von Vitamin C bei T-Lymphozyten, während die Befunde zu den Wirkungen einer Vitamin-C-Supplementation auf die Proliferation der T-Lymphozyten beim Menschen widersprüchlich ausfallen.
Das Chediak-Higashi-Syndrom*Es handelt sich hier um eine sehr seltene autosomal-rezessive Erbkrankheit, die phänotypisch in Form einer Reifestörung der Lysosomen in Leukozyten und Melanozyten imponiert. Dadurch kommt es zu Störungen der Hautpigmentierung und Infektabwehr. Symptome sind eine verminderte Pigmentierung (partieller Albinismus), silbrig-blondes Haar, Vergrößerung der Leber und Milz (Hepatosplenomegalie) und rezidivierende Infektionen. Die Lebenserwartung der Betroffenen ist stark vermindert; durch eine Knochenmarktransplantation ist die Erbkrankheit jedoch zu therapieren. Die Diagnose kann pränatal durch eine Blutuntersuchung oder auf Basis einer Haut-Haar-Biopsie gestellt werden. *in Anlehnung an Ströhle und Hahn [1] |
Funktionelle Bedeutung von Vitamin C für die Infektionsresistenz des Organismus
Eine Reihe von Untersuchungsergebnissen, die sowohl am Menschen als auch bei Vertretern anderer Tierspezies gemacht werden konnten, unterstreichen die funktionelle Bedeutung von Vitamin C für die Infektionsresistenz des Organismus. Folgende Beobachtungen sind dabei von Relevanz [1, 3, 26]:
Experimentell imponiert ein Vitamin-C-Defizit durch eine verminderte Infektionsresistenz gegenüber verschiedenen mikrobiellen Erregern wie Bakterien, darunter Mycobacterium tuberculosis und Rickettsia sowie Mykosen wie Candida albicans.
Eine verbesserte Vitamin-C-Versorgung steigert sowohl die Antikörper-Reaktion als auch die Aktivität der Neutrophilen und die Infektionsresistenz gegenüber Rabies-, Polio- und Parainfluenzaviren sowie gegen Salmonella gallinarum und E. coli. Tierexperimentell konnte unter Vitamin-C-Supplementierung zudem eine verminderte Mortalität bei Infektions- und Parasitenerkrankungen nachgewiesen werden. Konsistent hierzu ist auch die Beobachtung, wonach Vitamin C bei Herpes-simplex-, Rhino- und HIV-Viren virusstatische Effekte entfaltet. Die Ergebnisse einer – allerdings unkontrollierten – Interventionsstudie an HIV-positiven Personen deuten darauf hin, dass Megadosen an Vitamin C (50 – 200 g/d) das Risiko opportunistischer Infektionen verringern können [27].
Vitamin C wird bei Infektionserkrankungen rasch verbraucht; die Konzentration des Mikronährstoffs nimmt dabei im Plasma und in Leukozyten der betroffenen Organismen ab. Die Abnahme der Ascorbatkonzentration ist von einem Anstieg an Dehydroascorbat begleitet [3]. Während bei gesunden Personen 5 bis 10% des Vitamins in Form von Dehydroascorbat vorliegt, steigt dieser Anteil bei Patienten mit Meningokokken-Meningitis oder Pneumonie auf 65 bis 85% an [28]. Diese Beobachtung deutet darauf hin, dass Ascorbat bei Infektionen infolge der gesteigerten Bildung reaktiver Sauerstoffspezies (ROS) vermehrt oxidiert wird [26] und in dieser Situation auch die endogene enzymatische Reduktion an ihre funktionelle Grenze gelangt.
Immunologische Effekte von Vitamin C: Molekulare Mechanismen
Vor dem Hintergrund der eingangs des Artikels beschriebenen biochemischen Funktionen von Vitamin C lassen sich die immunmodulatorischen Effekte des Mikronährstoffs primär auf
- seine antioxidative Wirkung,
- seine Beteiligung an der de-novo-Carnitinsynthese,
- seine Relevanz für die Tetrahydrobiopterin-Synthese und auf seine
- Modulation des Eisenmetabolismus zurückführen [3].
Immunbiologische Relevanz der antioxidativen Kapazität von Vitamin C
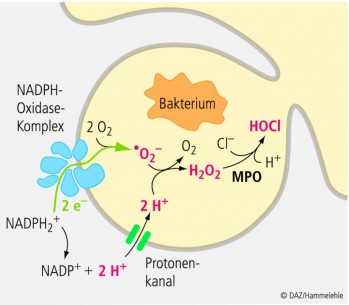
Aktivierte Phagozyten synthetisieren in einem NADPH-Oxidase-abhängigen Prozess (EC 1.6.3.1) hohe Mengen an reaktiven Superoxidanionenradikalen (O2 - .), die dann durch die Superoxiddismutase (SOD; EC 1.15.1.1) zu Wasserstoffperoxid (H2 O2) reduziert und/oder mit bereits synthetisiertem Wasserstoffperoxid unter Bildung der hochreaktiven Hydroxylradikale (OH.) reagieren. H2 O2 kann zudem unter dem Einfluss eines weiteren Enzyms, der Myeloperoxidase (EC 1.11.1.7), in Gegenwart von Chloridionen (Cl-) zu Hypochloridionen (OCl-) oxidiert werden. Funktional dienen die so erzeugten reaktiven Sauerstoffverbindungen der Abtötung von Bakterien (Abb. 4). Allerdings ist dieser für die Immunabwehr essentielle Prozess ausgesprochen janusköpfig, da auch die Phagozyten selbst durch die hochreaktiven Sauerstoffverbindungen geschädigt und in ihrer Funktion beeinträchtigt werden. Als effektives wasserlösliches Antioxidanz kommt Vitamin C daher eine zentrale Bedeutung für die Integrität der Phagozyten zu; überschüssig gebildete Sauerstoffradikale können von Vitamin C abgefangen und die Immunzellen so vor Schäden bewahrt werden. Insbesondere Makrophagen sind offenbar auf eine adäquate Vitamin-C-Versorgung angewiesen: Im Zuge der Phagozytose verbrauchen diese Vitamin C, das als eines der bedeutsamsten Antioxidantien in den Kompartimenten von Immunzellen lokalisiert ist [3]. Die zentrale Bedeutung der antioxidativen Eigenschaften von Vitamin C bei der immunologischen Abwehr lässt sich u. a. auf folgende Sachverhalte zurückführen [3]:
Das niedrige Redoxpotenzial von Ascorbat (282 mV) ermöglicht es dem Vitamin mit praktisch allen für biotische Systeme relevanten reaktiven Sauerstoff- und Stickstoffspezies zu interagieren und diese abzufangen. Auf diesem Weg wirkt Vitamin C als effektives Scavengermolekül unter anderem für Superoxidanionen (O2 - .), Hydroxyl- (OH.), Peroxidradikale (ROO-), Wasserstoffperoxid (H2 O2) und Peroxynitrit (ONOO).
Die Interaktion der Ascorbinsäure mit den oben genannten reaktiven Molekülen ist gekennzeichnet durch eine hohe Reaktionskonstante (> 105 mol/ l. s-1), womit die Neutralisation der Radikale effektiv, d. h. schnell und dauerhaft, erfolgen kann.
- Die oxidierte Form der Ascorbinsäure kann im Organismus enzymatisch auf effiziente Weise regeneriert werden. Durch diesen Recyclingprozess ist sichergestellt, dass ausreichende Mengen an biotisch aktivem Vitamin C zur Verfügung stehen.
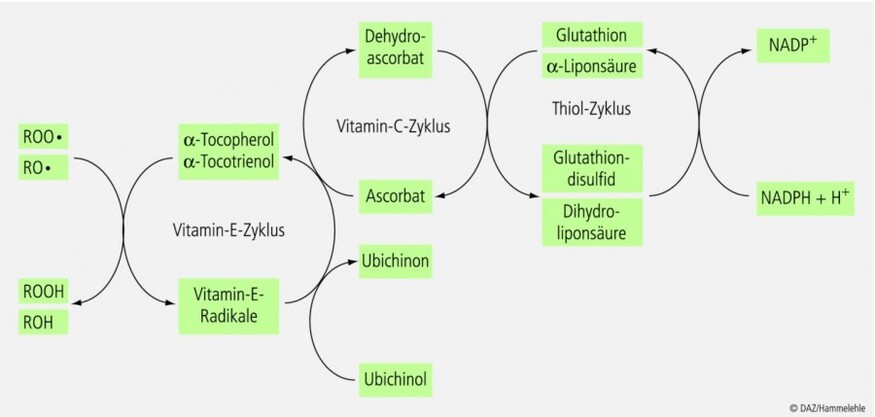
Vitamin C ist zentraler Metabolit im antioxidativen Netzwerk der Zelle. Von besonderer Bedeutung ist die Interaktion zwischen Vitamin C und Vitamin E (alpha-Tocopherol). Letzteres ist das wichtigste lipidlösliche Antioxidans des menschlichen Organismus und in der Lage, die Lipidperoxidation zu unterbrechen (Abb. 5). Das dabei gebildete alpha-Tocopheroxylradikal kann durch Vitamin C in die biotisch aktive Form alphaTocopherol reduziert werden. Darüber hinaus scheint Vitamin C eine "Sparwirkung" auf Vitamin E auszuüben, da es reaktive Sauerstoffspezies vor ihrer Reaktion mit Lipiden abfangen und damit die Lipidperoxidation verhindern kann.
Vitamin C induziert die Expression des Hämoxygenase-1/Ferritin-Systems. Über diesen Mechanismus wirkt Vitamin C auf indirektem Wege antioxidativ, da Ferritin intrazellulär freie Eisenionen binden und die eisenabhängige Synthese aggressiver Hydroxylradikale vermindern kann. Das Hämoxygenase-1-Produkt Billirubin fungiert im physiologischen Konzentrationsbereich zudem als Radikalfänger.
Immunbiologische Relevanz hinsichtlich der Carnitinsynthese von Vitamin C
Die Hydroxycarbonsäure Carnitin (β-Hydroxyγ-Trimethylaminobuttersäure) ist eine quaternäre Ammoniumverbindung. Carnitin wird endogen aus den Aminosäuren Lysin und Methionin in einem mehrstufigen Prozess synthetisiert. Dabei fungiert L-Methionin als Methylgruppendonator und proteingebundenes L-Lysin als Lieferant des übrigen Kohlenstoffskeletts. Als Cofaktoren werden bei diesem Syntheseprozess neben Eisen, Niacin und Vitamin B6 auch Vitamin C benötigt. Letzteres dient als Cofaktor zweier an der Carnitinsynthese beteiligter Enzyme, der
- β-Hydroxy-ε-N-Trimethyllysin-Hydroxylase (EC 1.14.11.8) und der
- γ-Butyrobetain-Hydroxylase (EC 1.14.11.1).
Erstere katalysiert die mitochondriale Hydroxylierung des durch Proteolyse aus zellulären Proteinen freigesetzten ε-N-Trimethyllysin, während Letztere die im Zytosol lokalisierte Hydroxylierung von γ-Butyrobetain einleitet.
Carnitin weist neben seiner Funktion als Biokarrier langkettiger Fettsäuren verschiedene immunogene Effekte auf. Dazu zählen [3]:
- Stimulation der Phagozytose und Chemotaxis von Granulozyten und Makrophagen,
- Stimulation der Immunglobulinsynthese (IgG),
- Stimulation der Akut-Phase-Antwort.
Vitamin C – Einfluss auf den Tetrahydrobiopterin- und NO-Stoffwechsel
Tetrahydrobiopterin (BH4) stellt im Stoffwechsel einen 1-Elektronendonator dar und ist Cofaktor der induzierbaren Stickstoffmonoxid-Synthase (NOS; EC 1.14.13.39), die über mehrere Stufen Arginin zu Citrullin und NO oxidiert. BH4 reagiert leicht mit reaktiven Sauerstoffspezies, insbesondere Peroxynitrit, wodurch BH4 zu einem Trihydrobiopterinradikal oxidiert, inaktiviert und damit der NO-Synthese entzogen wird. Ascorbinsäure steigert die NO-Synthese, indem es Trihydrobiopterinradikale zu biotisch aktivem BH4 reduziert und/oder BH4 vor Oxidation schützt. Damit kommt Vitamin C eine entscheidende Bedeutung für die NO-Verfügbarkeit der Zelle zu [3].
Die immunologische Relevanz dieses Aspektes ist auf die Tatsache zurückzuführen, dass NO eine Schlüsselsubstanz für die zelluläre Immunität darstellt. So synthetisieren zytokin-aktivierte Makrophagen vermehrt NO, das einerseits mikrobizide Effekte entfaltet und andererseits die Funktion von T-Lymphozyten moduliert (Inhibierung der T-Lymphozytenmitogenese und Induktion der Transformation von Th1- zu Th2-Helferzellen [3].
Zu den weiteren Wirkmechanismen, über die Vitamin C Einfluss auf die Aktivität der zellulären Immunabwehr nimmt, zählen [3]:
- Synthese von Interferon γ, ein für die Aktivität der natürlichen Killerzellen relevantes und die Virusreplikation blockendes Glykoprotein;
- Synthese von cyclischen Nukleotiden und von RNA in Immunzellen.
Effekte von Vitamin C bei Erkältungskrankheiten
Basierend auf der bereits erwähnten, 1970 von Linus Pauling (1901 – 1994) verfassten Monographie "Vitamin C and the Common Cold", wurde immer wieder diskutiert, ob die Gabe hoher Mengen an Vitamin C sowohl die Anfälligkeit als auch die Dauer und Schwere von Erkältungskrankheiten reduzieren kann. Zwischenzeitlich liegt eine Vielzahl an Interventionsstudien vor, die die postulierten prophylaktischen Wirkungen einer Vitamin-C-Supplementierung zum Untersuchungsgegenstand hatten. Die darauf basierenden Nachauswertungen und Metaanalysen zeigen, dass die Gabe von Vitamin C (≥ 200 mg/d über mehrer Wochen bis mehrere Jahre) folgende Effekte entfaltet [1]:
Im Hinblick auf die Krankheitshäufigkeit ist die prophylaktische Supplementierung von Vitamin C in der Normalbevölkerung generell ohne klinischen Nutzen, wie die Auswertung von 30 Interventionsstudien mit 11.350 Teilnehmern ergab. Das relative Risiko (RR) an einer Erkältung zu erkranken, schwankte in den Verumgruppen der Einzelstudien zwischen 0,39 und 1,36; der gepoolte RR-Wert aller Studien betrug 0,96 (95% CI 0,92 – 1,00). Dagegen zeigt die Subgruppenanalyse von sechs Interventionsstudien, dass Personen, die besonderen Stresssituationen mit erhöhter Infektanfälligkeit ausgesetzt sind (z. B. Marathonläufer), von einer prophylaktischen Vitamin-C-Einnahme profitieren (RR 0,50; 95% CI 0,38 – 0,66) [29].
Im Hinblick auf die Krankheitsdauer ist die prophylaktische Supplementierung von Vitamin C sowohl bei Erwachsenen (18 Studien mit 7242 Krankheitsfällen) als auch bei Kindern (zwölf Studien mit 2434 Krankheitsfällen) mit einem Benefit verbunden. Für Erwachsene ergab die gepoolte Auswertung aller Studien eine Reduktion der Krankheitsdauer von 8% (95% CI -13,8 bis -2,96); bei Kindern war der Effekt noch deutlicher (Krankheitsdauerreduktion 13,6%; 95% CI -21,6 bis -5,6) [29].
Im Hinblick auf die Krankheitsschwere ist die prophylaktische Supplementierung von Vitamin C in der Normalbevölkerung generell ohne klinischen Nutzen, wie die Auswertung von acht Interventionsstudien mit 1979 Teilnehmern ergab (gepoolter Effekt 0,13% 95% CI -0,21 bis -0,04).
Unabhängig von der Frage, ob und inwieweit die prophylaktische Einnahme von Vitamin-C-Supplementen von einem klinischen Benefit begleitet wird, ist die Frage von Interesse, welche therapeutischen Effekte von Vitamin C bei manifesten Erkältungskrankheiten entfaltet. Auch zu dieser Fragestellung liegt eine Vielzahl an Untersuchungen vor. Eine darauf basierende Metaanalyse erbrachte folgendes Kernergebnis: Megadosen an Vitamin C (im Grammbereich) zeigen weder im Hinblick auf die Dauer (gepoolter Effekt -2,54, CI -10,09 bis -5,02) noch im Hinblick auf die Schwere (gepoolter Effekt -0,007 95% CI -0,16 bis 0,02) der Erkältungssymptome einen klinisch relevanten Nutzen [29].
Teil 2 des Beitrags "Immunbiologische Effekte von Mikronährstoffen" folgt in DAZ 2011, Nr. 12.
Literatur
[1] Ströhle A, Hahn A. Vitamin C und Immunfunktion. Med Monatsschr Pharm. 2009; 32:49 – 54
[2] Wintergerst ES, Maggini S, Hornig DH. Contribution of selected vitamins and trace elements to immune function. Ann Nutr Metab 2007; 51:301 – 23
[3] Ströhle A, Wolters M, Hahn A. Micronutrients at the interface between inflammation and infection ascorbic Acid and calciferol: part 1, general overview with a focus on ascorbic Acid. Inflamm Allergy Drug Targets. 2011; 10:54 – 63
[4] Ströhle A, Wolters M, Hahn A. Micronutrients at the interface between inflammation and infection ascorbic Acid and calciferol. Part 2: calciferol and the significance of nutrient supplements. Inflamm Allergy Drug Targets. 2011;10:6474
[5] Ströhle A, Hahn A. [Importance of micronutrients for immunity--preventive and therapeutic aspects]. MMW Fortschr Med. 2009;151 Suppl 3:133 – 41
[6] van Lettow M, Fawzi WW, Semba RD. Triple trouble: the role of malnutrition in tuberculosis and human immunodeficiency virus co-infection. Nutr Rev 2003; 61:81 – 90
[7] Semba RD, Tang AM. Micronutrients and the pathogenesis of human immunodeficiency virus infection. Br J Nutr 1999; 81:181 – 9
[8] Colecraft E. HIV/AIDS: nutritional implications and impact on human development. Proc Nutr Soc. 2008; 67:109 – 13
[9] Adam O. Dietary therapy for inflammatory rheumatic diseases. Dtsch Med Wochenschr. 2009; 134:1759 – 63
[10] Adam O, Fasse S, Ditrich O. Diet in rheumatic disease. Z Rheumatol 2009; 68:549 – 58
[11] Hahn A, Ströhle A, Wolters M. Ernährung bei Erkrankungen des rheumatischen Formenkreises. Med Monatsschr Pharm 2007; 30:138 – 46
[12] Meydani SN, Wu D. Nutrition and age-associated inflammation: implications for disease prevention. JPEN J Parenter Enteral Nutr 2008; 32:626 – 9
[13] Lau FC, Shukitt-Hale B, Joseph JA. Nutritional intervention in brain aging: reducing the effects of inflammation and oxidative stress. Subcell Biochem 2007; 42:299 – 318
[14] Calder PC, Albers R, Antoine JM, Blum S, Bourdet-Sicard R, Ferns GA, Folkerts G, Friedmann PS, Frost GS, Guarner F, Løvik M, Macfarlane S, Meyer PD, M‘Rabet L, Serafini M, van Eden W, van Loo J, Vas Dias W, Vidry S, Winklhofer-Roob BM, Zhao J. Inflammatory disease processes and interactions with nutrition. Br J Nutr 2009; 101 Suppl 1:S1 – 45
[15] Pattison DJ, Winyard PG. Dietary antioxidants in inflammatory arthritis: do they have any role in etiology or therapy? Nat Clin Pract Rheumatol 2008; 4:590 – 6
[16] Pauling L. Vitamin C and the Common Cold. Freeman, San Francisco 1970
[17] Padayatty SJ, Levine M. New insights into the physiology and pharmacology of vitamin C. CMAJ. 2001; 164:353-5
[18] Heller R. Ascorbat. In: Dunkelberg H, Gebel T, Hartwig A (Hrsg.): Handbuch der Lebensmitteltoxikologie. Belastungen, Wirkungen, Lebensmittelsicherheit, Hygiene. Weinheim: Wiley-VCH, 2007; 2135 – 62
[19] Padh H. Cellular functions of ascorbic acid. Biochem Cell Biol 1990; 68:1166 – 7390
[20] Kaelin WG. Proline hydroxylation and gene expression. Annu Rev Biochem 74: 115-28, 2005.
[21] Hausinnger RP. Fell/alpha-ketoglutarate-dependent hydroxylases and related enzymes. Crit Rev Biochem Mol Biol 39(1): 21 – 68, 2004.
[22] Siddiq A, Aminova LR, Ratan RR. Prolyl 4-hydroxylase activity-responsive transription factors: from hydroxylation to gene expression and neuroprotection. Front Biosci 13: 2875 – 87, 2008.
[23] Levine M, Conry-Cantilena C, Wang Y, Welch RW, Washko PW, Dhariwal KR, Park JB, Lazarev A, Graumlich JF, King J, Cantilena LR. Vitamin C pharmacokinetics in healthy volunteers: evidence for a recommended dietary allowance. Proc Natl Acad Sci USA 1996; 93:3704 – 9.
[24] Jacob RA, Kelley DS, Pianalto FS, et al. Immunocompetence and oxidant defense ring corbate depletion of healthy men. Am J Clin Nutr 1991; 54:1302 – 9.
[25] Webb AL, Villamor E. Update: effects of antioxidant and non-antioxidant vitamin supplementation on immune function. Nutr Rev 2007; 65:181 – 217.
[26] Hemilä H. Vitamin C and Infectious Diseases. In: Packer L, Fuchs J (eds.): Vitamin C in Health and Disease. Marcel Dekker, New York 1997a; 471 – 503.
[27] Cathcart RF 3rd. Vitamin C in the treatment of acquired immune deficiency syndrome (AIDS). Med Hypotheses 1984; 14:423 – 33.
[28] Chakrabarti B, Banerjee S. Dehydroascorbic acid level in blood of patients suffering from various infectious diseases. Proc Soc Exp Biol Med 1955; 88:581 – 3.
[29] Douglas RM, Hemilä H, Chalker E, Treacy B. Vitamin C for preventing and treating the common cold. Cochrane Database Syst Rev 2007; 18;(3):CD000980
Autoren
Dr. Alexander Ströhle*,
Dr. Maike Wolters,
Prof. Dr. oec. troph. Andreas Hahn
* Anschrift des Korrespodenzautors:
Dr. Alexander Ströhle
Institut für Ernährungsphysiologie und Humanernährung,
Zentrum für angewandte Chemie,
Leibniz Universität Hannover
Am Kleinen Felde 30
30167 Hannover
Fax: +49-511-762-5729
E-Mail: stroehle@nutrition.uni-hannover.de



0 Kommentare
Das Kommentieren ist aktuell nicht möglich.