
















- DAZ.online
- DAZ / AZ
- DAZ 48/2011
- HIV-Impfstoffentwicklung
Schwerpunkt HIV und AIDS
HIV-Impfstoffentwicklung
1988 wurde mit der National Cooperative Vaccine Development Groups for AIDS (NCVDG) ein erstes Konsortium aus Regierung, Wissenschaft und Industrie gegründet, in dem man sich gezielt mit der Impfstoffentwicklung auseinandersetzte. In den 1990er Jahren organisierten sich die HIV-Impfstoffforscher in zahlreichen weiteren Konsortien, wie der International AIDS Vaccine Initiative (IAVI), der EuroVacc, der Collaboration for AIDS Vaccine Discovery (CAVD), dem Center for HIV/AIDS Vaccine Immunology (CHAVI) und der South African AIDS Vaccine Initiative (SAAVI). Obwohl also seit Jahren wirklich zahlreiche namhafte Wissenschaftler an dem dringenden Problem arbeiten und weltweit jährlich mehr als 950 Millionen US-$ in die HIV-Impfstoffforschung fließen, ist es noch immer nicht gelungen, eine wirksame Vakzine zu entwickeln.
Epidemiologie
Das humane Immundefizienzvirus HIV gehört zu den Retroviren und hier genauer zu den Lentiviren (siehe Infokasten "Aufbau und Lebenszyklus von HIV"). Zwei Vertreter sind bekannt: HIV-1 und HIV-2. Sie nutzen die gleichen Übertragungswege, und beide können die Erkrankung AIDS auslösen. Allerdings ist HIV-2 im Wesentlichen auf Gebiete in Westafrika beschränkt und "nur" ca. 1 bis 2 Millionen Menschen sind weltweit mit HIV-2 infiziert. Zudem sind HIV-2-Infizierte zu 80% Langzeitüberlebende, und nur eine Minderheit entwickelt tatsächlich das Vollbild AIDS, wobei die Krankheit dann auch nur relativ langsam voranschreitet.
Anders sieht es mit HIV-1 aus: Dieses Virus ist für den Großteil der HIV-Infektionen verantwortlich; nach wie vor leben weltweit ca. 35 Millionen Menschen mit HIV-1. Immerhin sind aber die Zahlen der jährlich dazukommenden Neuinfektionen leicht rückläufig. Sie lagen laut UNAIDS Report on the Global AIDS Epidemic 2009 bei ca. 2,6 Millionen, während es beim Höchststand im Jahr 1997 3,2 Millionen Neuinfektionen waren. Auch die Zahl der AIDS-bedingten Todesfälle ist mit 1,8 Millionen im Jahr 2009 im Vergleich zum Maximum von 2,1 Millionen im Jahr 2004 leicht rückläufig.
Aktuelle Schätzungen aus dem Robert-Koch-Institut zeigen, dass bis Ende 2011 in Deutschland 73.000 HIV-positive Menschen leben werden, die Zahl der Neuinfektionen im Jahr 2011 wird auf 2700 geschätzt. Im Zeitraum 1982 bis 2010 wurden insgesamt 28.027 AIDS-Fälle registriert. 14.178 der Erkrankten sind mittlerweile verstorben.
Nach wie vor wird eine HIV-Infektion und AIDS als die wichtigste Infektionserkrankung wahrgenommen, obwohl mittlerweile – zumindest in den Industrieländern – sehr effiziente antiretrovirale Medikamente zur Verfügung stehen, die eine HIV-Infektion so gut in Schach halten, dass sie von einer akut tödlichen Gefahr zu einer chronischen Erkrankung umdefiniert wurde. Aber die Betroffenen sind infiziert, und sie können weitere Menschen infizieren. Außerdem stehen diese Wirkstoffe gerade in den Hochprävalenzländern nicht oder nicht im ausreichenden Umfang zur Verfügung. Deshalb ist ein Impfstoff, der nachhaltig vor einer Infektion schützt, nach wie vor eine dringlichst benötigte Waffe gegen HIV-Infektionen und AIDS.
Der Welt-AIDS-Tag am 1. Dezember 2011 gibt Anlass, wieder einmal den Status quo und die Zukunftsaussichten in der HIV-Impfstoffforschung zu reflektieren.
Aufbau und Lebenszyklus von HIVHIV gehört zu den Lentiviren aus der Familie der Retroviridae. Eine Infektion mit einem derartigen, "langsamen" Virus resultiert in einer relativ langsam fortschreitenden Erkrankung, in diesem Fall von AIDS, dem Acquired Immunodeficiency Syndrome. HIV-1 ist ein umhülltes Virion mit einem Durchmesser von 80 bis 100 nm. Als Retrovirus hat es eine einzelsträngige RNA als Genom, die in zwei Kopien im Viruspartikel vorliegt. Die ca. 9700 Basen lange RNA weist eine 5’-Cap-Struktur sowie eine 3’-Polyadenylierung auf und ähnelt dadurch einer mRNA aus eukaryontischen Zellen. Mit der RNA sind im Viruspartikel Nucleocapsidproteine (p7) assoziiert. Innerhalb des konischen Capsids von HIV befinden sich neben dem Nucleocapsid noch die viralen Proteine Protease, Integrase und Reverse Transkriptase. Ein Link-Protein (p6) bildet die Verbindung zwischen Capsid und Hüllmembran. In der Membran, die sich von der Cytoplasmamembran der Wirtszelle ableitet, befinden sich die viralen, trimeren Envelope-Proteine (Env), die zunächst als gp160-Vorläuferglykoproteine gebildet und dann in die Untereinheiten gp120 und gp41 geschnitten werden. Die Innenseite der Hüllmembran wird von Matrixprotein (p17) ausgekleidet (Abbildung Aufbau HI-Virus). Für die Infektion einer Wirtszelle bindet gp120 an das CD4-Molekül und einen weiteren Co-Rezeptor in der Plasmamembran der Wirtszelle (Abbildung Entwicklungszyklus des HI-Virus). Durch Konformationsänderung des Glykoproteinkomplexes kommt es zur Fusion zwischen der Virus-Hüllmembran und der Zellmembran der Wirtszelle, wodurch das virale Capsid mit der RNA in die Zelle gelangt. Im Cytoplasma der Wirtszelle schreibt die mitgebrachte, virale Reverse Transkriptase das RNA-Genom in doppelsträngige DNA um. Anschließend kommt die virale Integrase zum Zug und integriert das umgeschriebene Virusgenom an einer beliebigen Stelle in die DNA der Wirtszelle. Hier verbleibt das virale Genom, wird bei jeder Zellteilung an die Tochterzellen weitergegeben und über die zelluläre RNA-Polymerase II transkribiert. Die entstehenden RNA-Moleküle dienen einerseits als genomische RNA für neue Virus-Partikel, andererseits als mRNA für die Bildung der viralen Proteine. Die extreme Heterogenität von HIV ist das Resultat des schnellen Virus-Turnovers von ca. 2,6 Tagen pro Replikationszyklus, der hohen Viruslast von ca. 1010 gebildeten Viruspartikeln pro Tag und der fehlerhaft arbeitenden Reversen Transkriptase, die bei der DNA-Synthese statistisch 3 Fehler pro 100.000 Basen macht. In einem infizierten Patienten befindet sich schließlich ein Schwarm von genotypisch ähnlichen Virus-Klonen, die als Quasi-Spezies bezeichnet werden. |
Sehr viele verschiedene HIV-1-Varianten
Eines der ganz großen Probleme bei der Impfstoffentwicklung gegen HIV-1 ist die extreme strukturelle Hypervariabilität des Retrovirus (siehe Tab. 1 und Infokasten). Aufgrund der hohen Sequenzunterschiede von mehr als 30% teilt man HIV-1 mittlerweile in drei Gruppen ein, die als
M (main-Gruppe),
O (outlier-Gruppe) und
N (Non-M/Non-O-Gruppe)
bezeichnet werden. Viren der Gruppe M sind für den Großteil der Infektionen und AIDS-Erkrankungen verantwortlich und werden nochmals in die Subtypen (engl. clades) A bis D, F bis H, J und K unterteilt, wobei sich die Subtypen A und F nochmal jeweils aus den sogenannten Sub-Subtypen A1, A2 und A3 bzw. F1 und F2 zusammensetzen.
Tab. 1: Wissenschaftliche Herausforderungen in der Entwicklung eines Impfstoffes gegen AIDS (AIDS Vaccine Blueprint 2008) | |
HIV ist ein Retrovirus |
|
HIV induziert keine protektive Immunität |
|
HIV ist hypervariabel |
|
HIV entzieht sich der Erkennung durch das Immunsystem |
|
HIV infiziert nur Menschen |
|
HIV wird bei Sexualkontakt übertragen |
|
Die Verteilung der Subtypen ist weltweit ungleich. Der Subtyp B ist vor allem in den Industrieländern, in Lateinamerika und in der Karibik vorherrschend, während in Zentralafrika vor allem die Subtypen A und D vorkommen. In Südafrika, Teilen Ostafrikas und Indien ist hingegen der Subtyp C für die meisten Infektionen verantwortlich.
Die Sequenzähnlichkeit zwischen Viren verschiedener Subtypen liegt zwischen 70 und 90%. Vor allem im env-Gen unterscheiden sich jedoch die einzelnen Subtypen zu ca. 30%. Bei der Doppelinfektion einer Zelle mit mindestens zwei unterschiedlichen Virus-Subtypen kommt es sehr häufig zu Rekombinationen zwischen den Viren, und es entstehen dabei sogenannte zirkulierende rekombinante Formen (circulating recombinant forms, CRF). CRFs sind für ca. 10 bis 20% aller Neuinfektionen verantwortlich. Mittlerweile finden sich 51 CFRs in der HIV Sequence Database am Los Alamos National Laboratory, von denen die Hauptformen CRF01_AE, CRF02_AG, CRF07,08_BC und CRF_BF in Asien, Westafrika, China und Südamerika vorkommen.
Zusätzlich existieren noch unique recombinant forms (URF) von HIV, die sehr unterschiedliche Rekombinations-Bruchpunkte aufweisen und nur eine begrenzte Verbreitung in der Bevölkerung haben. In Gegenden, in denen verschiedene HIV-Subtypen zirkulieren, wie z. B. Uganda und Kenia, sind URFs an über 30% der Infektionen beteiligt.
Grafik: Zündorf
Der steinige Weg der HIV-Impfstoffentwicklung
Die Geschichte der HIV-Impfstoffentwicklung (Abb. 1) lässt sich in drei Etappen einteilen. Die erste Etappe umspannte die Jahre 1983 bis 1994, die zweite ging von 1995 bis 2007 und die dritte begann 2008 und dauert noch an.
Nachdem im Jahr 1981 in den USA ein Impfstoff gegen das Hepatitis-B-Virus zugelassen worden war, war man zunächst sehr zuversichtlich, dass man die gewonnenen Erfahrungen nutzen könne, um sehr schnell einen analogen Impfstoff gegen HIV zu entwickeln. Für HIV kam als potenzielles Antigen nur das Env-Protein und hier vor allem der gp120-Anteil infrage (siehe Infokasten). Dieses Protein ist auf der Virusoberfläche exponiert, und über dieses Protein dockt das Virus an die Wirtszelle an – eigentlich ideale Voraussetzungen für die Induktion neutralisierender Antikörper. Parallel dazu wurde eine Vakzine mit attenuierten Affenviren in Makaken getestet. Bei beiden Ansätzen wurden zwar neutralisierende Antikörper nachgewiesen, allerdings schützten sie nur vor HIV-Laborstämmen, nicht jedoch vor Wild isolaten. Der Ansatz mit attenuierten Viren wurde nicht weiter verfolgt, weil die Gefahr einer Rückmutation oder Rekombination zu infektiösen HI-Viren als deutlich zu groß eingeschätzt wurde. 1994 wurden auch die Arbeiten an Env-basierten Protein- und Peptid-Impfstoffen wegen des fehlenden Erfolgs eingestellt.
Die nächste Runde der Impfstoff entwicklung ging einher mit einer konzeptionellen Neupositionierung. Bei den neuen Vakzinen setzte man nicht mehr nur auf neutralisierende Antikörper mit einem breiten Wirkspektrum, sondern zusätzlich auch auf die Induktion einer Zell-vermittelten Immunität. Die Ratio für diesen erweiterten Ansatz lag darin, dass neutralisierende Antikörper zwar freie Viruspartikel abfangen können, dass aber vor allem zytotoxische T-Zellen infizierte Wirtszellen aufspüren und möglichst eliminieren sollten. Um dieses Ziel zu erreichen wurden in erster Linie lebende virale Vektoren, beispielsweise mit HIV-Genen modifizierte Pocken- oder Adenoviren, als Vakzine verwendet. Zudem wurden viele dieser Vektoren in einem sogenannten Prime-Boost-Ansatz zusammen mit DNA-Vektoren eingesetzt, die ebenfalls HIV-Genombereiche enthielten. Und schließlich wurden die Impfstoffe auch noch mit gereinigtem gp120-Protein kombiniert. Dahinter stand das Konzept, zunächst mit der DNA zu immunisieren (Prime) und dann mit viralen Vektoren die initiale Impfwirkung zu verstärken (Boost).
Aufsehen erregte in dieser zweiten Etappe die STEP-Studie, bei der replikations-inkompetente Adenoviren des Serotyps 5 (Ad5) verwendet wurden, die für die relativ konservierten HIV-Proteine Gag, Pol und Nef codierten (siehe Infokasten). Dieser Impfstoff erwies sich in Tests an nicht-humanen Primaten zunächst als sehr vielversprechend. Allerdings traten bei der eigentlichen STEP-Studie dann sogar mehr HIV-Infektionen in der Gruppe der Geimpften auf als in der Placebo-Gruppe, weshalb die Studie abgebrochen wurde.
Mit diesem erneuten Misserfolg endete quasi die zweite Welle der HIV-Impfstoffentwicklung.
Eine Phase-III-Studie aus dieser Zeit gibt jedoch nach wie vor Grund zur Hoffnung: Im Oktober 2003 startete eine groß angelegte Studie in Thailand (RV144) mit über 16.000 Probanden, bei der rekombinante Kanarienpockenviren als Prime-Injektionen verwendet wurden, die für die HIV-Proteine Gag, Pol und Env des Virus-Subtyps E codierten (ALVAC). Die anschließenden Booster-Impfungen enthielten rekombinantes Glykoprotein 120 der Virus-Subtypen B und E (AIDSVAX). Bei dieser Studie zeigte sich eine leichte Schutzwirkung der Vakzine von 31%. Allerdings hatte die Immunisierung keinerlei Effekt auf die Viruslast oder die CD4+-T-Zellzahl. Dennoch wurde hier erstmals ein gewisser Schutz beobachtet, dessen Mechanismus nach wie vor analysiert wird.
Auch die Ergebnisse der STEP-Studie und der STEP-ähnlichen Phambili-Studie in Südafrika werden sehr sorgfältig dahingehend ausgewertet, ob sich Patienten-Subgruppen definieren lassen, die besser ansprechen als andere, um daraus für zukünftige Impfstoffentwicklungen zu lernen.
Mit diesen Erfahrungen startete die dritte Welle der HIV-Impfstoffentwicklung, die nach wie vor auf die Induktion sowohl einer Zell-Immunität als auch von neutralisierenden Antikörpern setzt. Allerdings werden nicht mehr die groß angelegten Studien initiiert. Vielmehr werden mehrere kleine Studien mit einer begrenzten und definierten Probandenauswahl durchgeführt. So können schneller neue Erkenntnisse zu den gewählten Immunisierungsstrategien und den daraus resultierenden Immunantworten gewonnen und für mögliche neue Ansätze umgesetzt werden.
Derzeit laufen laut International AIDS Vaccine Initiative ungefähr 30 Impfstoffstudien vor allem in den frühen Phasen I und II, bei denen überwiegend Kombinationen von verschiedenen Antigenen einsetzt werden.
Warum es so schwierig ist, einen guten HIV-Impfstoff zu etablieren
Margaret Johnston und Anthony Fauci formulierten in einem kürzlich im New England Journal of Medicine erschienenen Kommentar das Problem sehr anschaulich: Erfolgreiche Impfstoffe simulieren einfach nur die normale Immunantwort. Infiziert man sich beispielsweise mit Masern, Mumps oder Windpocken, reagiert in den meisten Fällen das Immunsystem des Betroffenen sehr schnell, und es kommt zur Elimination des Pathogens durch neutralisierende Antikörper und zytotoxische T-Zellen. Die für diese Krankheiten entwickelten, bereits erfolgreich eingesetzten Impfstoffe – in den genannten Beispielen attenuierte Viren – induzieren dann nur eine ganz normale Immunantwort wie im Falle einer tatsächlichen Infektion. Sie stimulieren also B- und T-Zellen, ohne dass es jedoch zum Ausbruch der eigentlichen Erkrankung kommt.
Ganz anders sieht es dagegen bei HIV aus: Die Infizierten zeigen kaum eine natürliche Immunantwort, die in der Lage wäre, das Virus zu eliminieren oder wenigstens in Schach zu halten. Will man daher eine effizient schützende HIV-Vakzine entwickeln, muss man versuchen, die Immunantwort gegen HIV zu verbessern. Es muss das Immunsystem in einer Weise stimuliert werden, wie das natürlicherweise gar nicht der Fall ist.
Grafik: Zündorf
Von HIV-Infizierten lernen
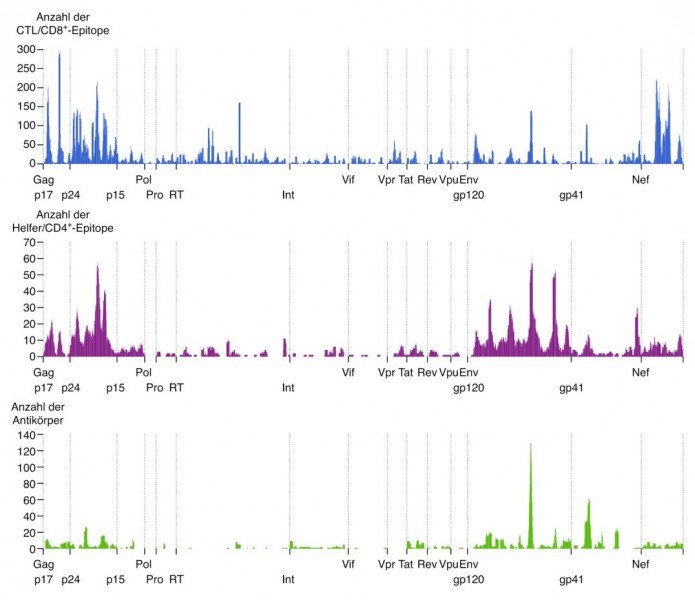
Aber wie reagiert eigentlich das Immunsystem bei einer Infektion mit HIV? Mittlerweile werden nicht nur Teilnehmer an den Impfstudien, sondern auch viele HIV-Infizierte selbst genauer hinsichtlich ihrer Immunantwort untersucht. All die gefundenen B-Zell- und T-Zellantworten wurden gesammelt und in der HIV Immunology Database katalogisiert. Eine Vielzahl an spezifischen Epitopen sind inzwischen identifiziert und charakterisiert, wobei auffällt, dass sich Epitope für zytotoxische T-Zellen vor allem im Matrix- und Capsidprotein sowie im Nef-Protein finden (Abb. 2). T-Helferzellen erkennen aktivierende Epitope ebenfalls im Matrix- und im Capsidprotein, zusätzlich aber auch im Glykoprotein 120. Erwartungsgemäß liegen die effizientesten Antikörper-Epitope vor allem in dem nach außen gerichteten Env-Protein, also sowohl im gp120 als auch im gp41.
Verfolgt man den Verlauf der Immunantwort nach der HIV-Übertragung, dann zeigt sich, dass bereits innerhalb weniger Tage Akute-Phase-Proteine aktiviert werden. Einige Tage später findet man erhöhte Zytokinkonzentrationen im Plasma, und auch die erste Antikörperantwort gegen das Oberflächen-Glykoprotein Env manifestiert sich innerhalb der ersten zwei Wochen nach Infektion. Allerdings haben diese Antikörper keine neutralisierende Wirkung (Abb. 3). Ungefähr 10 bis 30% der chronisch HIV-Infizierten bilden jedoch nach einigen Monaten bis Jahren Antikörper, die verschiedene HIV-Varianten binden und zu einem gewissen Ausmaß auch neutralisieren können. Nur bei ca. 2% der HIV- Infizierten finden sich sogenannte broadly neutralizing antibodies (bNAbs), die sehr effizient gegen ein breites Spektrum an HIV-Stämmen wirksam sind. Behandelt man nicht-humane Primaten mit derartigen Antikörpern, werden die Affen durch diese passive Immunisierung vor einer HIV-Infektion geschützt.
Grafik: Zündorf
Genau diese bNAbs sind es, die mittlerweile in den Fokus der Impfstoffforscher gerückt sind. Alle bisher genauer charakterisierten bNAbs zeigen entweder extrem hohe somatische Mutationsraten oder eine sehr lange dritte hochvariable Region (CDR3) in der schweren Antikörperkette. Typischerweise akkumulieren Antikörper während ihres Entwicklungs- und Reifungsprozesses 5 bis 15% Mutationen in der Aminosäuresequenz der variablen Domänen. bNAbs akkumulieren jedoch bis zu 30% und mehr Mutationen (Tab. 2).
Rein theoretisch wäre es denkbar, einen oder auch mehrere dieser bNAbs rekombinant herzustellen und als passive Immunisierung zu verwenden, ähnlich wie Synagis® bei einer drohenden RSV-Infektion. Aber im Gegensatz zu Synagis®, das nur in einem recht begrenzten Zeitraum einer recht überschaubaren Patientenpopulation verabreicht wird, müsste eine passive HIV-Immunisierung Milliarden Menschen ein Leben lang gegeben werden, was natürlich absolut undenkbar ist. Einzig in der Postexpositionsprophylaxe, z. B. bei medizinischem Personal nach Kontakt mit kontaminiertem Material, oder aber um ein ungeborenes Kind vor einer Ansteckung durch die HIV-positive Mutter zu schützen, wäre eine derartige passive Immunisierung sinnvoll anzu wenden.
Gelänge es jedoch, das Immunsystem durch eine aktive Immunisierung dazu zu bringen, solche bNAbs direkt zu bilden und nicht erst nach jahrelanger chronischer Infektion und das auch nur bei einigen wenigen Individuen, dann sollte es eigentlich möglich sein, HI-Viren frühzeitig abzufangen und zu neutralisieren.
Alle bNAbs binden an das Env-Protein auf den HI-Viruspartikeln, wobei sich ihre Bindungs-Spezifität in vier Gruppen einteilen lässt (Tab. 2):
die Membran-nahe externe Region von gp41 (membrane proximal external region, MPER),
Kohlenhydratgruppen auf gp120
die Bindungsstelle für CD4,
sowie der quarternäre V2/V3-Loop in gp120.
Tab. 2: Charakteristika repräsentativer bNAbs (mod. nach Verkoczy, et al. Curr Opin Immunol, 2011) | ||||
bNAba
|
Env Epitop-Spezifität |
Neutralisationb
|
% VH
- Mutationenc |
|
Breite |
Potenz |
|||
2F5 |
gp41 MPER |
60 |
2,3 |
15,2 |
4E10 |
gp41 MPER |
98 |
3,2 |
15,6 |
M66.6 |
gp41 MPER |
25 |
18,0 |
9,3 |
2G12 |
gp120 Glycan-Struktur |
32 |
2,4 |
31,7 |
1b12 |
gp120 CD4-Bindestelle |
41 |
1,8 |
13,1 |
VRC01, VRC02 |
gp120 CD4-Bindestelle |
91 |
0,34, 0,32 |
32,1 |
VRC03 |
gp120 CD4-Bindestelle |
57 |
0,45 |
30,2 |
HJ16 |
gp120 CD4 DMR Region |
36 |
8,0 |
14,6 |
PG9, PG16 |
gp120 quarternäre V2, V3 |
79, 73 |
0,22, 0,15 |
16,7, 20,5 |
CH01-CH04 |
gp120 quarternäre V2, V3 |
35 – 45 |
0,46 – 1,1 |
11,5 – 14,3 |
| a bNAbs in einer Reihe sind somatische Varianten des gleichen Klons b "Breite" ist der Anteil der HIV-Primärisolate, die mit einem IC50 < 50µg/mL neutralisiert werden; "Potenz" ist definiert als der IC50-Mittelwert [µg/mL] unter denjenigen Virus-Varianten, die mit einem IC50-Wert < 50µg/mL neutralisiert werden c geschätzt anhand der Aminosäuresequenz des Antikörpers im Vergleich zu den ursprünglichen V(D)J-Rearrangements der B-Zelle | ||||
Das optimale Immunogen
Die antigenen Strukturen von Env, an die die bNAbs binden, sind mittlerweile genau charakterisiert. Kann man aber dieses Wissen wirklich nutzen, um auf dieser Basis einen optimalen Impfstoff herzustellen?
Die Zuckerstrukturen, die von einem der bNAbs erkannt werden, fallen schon mal als Antigen weg: Sie sind zu ähnlich mit den Kohlenhydraten, die sich auf vielen Zellen des infizierten Wirts befinden. Somit könnten die gebildeten Antikörper kreuzreagieren und zu einer ungewollten Eliminierung von gesunden Körperzellen führen.
Drei der bisher charakterisierten bNAbs binden an die Membran-nahe externe Region von gp41 (membrane proximal external region, MPER). MPER ist eine Tryptophan-reiche Region von gp41 und stellt als relativ konservierte Domäne eine interessante immunologische Zielstruktur dar. Die genauere Analyse der Antikörper/Antigen-Interaktion zeigte, dass die Hüllmembran des Virus eine entscheidende Rolle bei der Bindung des Antikörpers hat. Auf der Basis dieser Erkenntnisse wurden bereits verschiedene Antigene entworfen, die die Peptid/Membran-Epitope simulieren. Allerdings konnten mit diesen Antigenen bisher keine breit wirksamen neutralisierenden Antikörper induziert werden.
Große Hoffnungen liegen nach wie vor auf gp120 und hier vor allem auf der Bindungsstelle, über die der Kontakt zum CD4-Protein auf der Wirtsstelle stattfindet. Diese Region ist zum einen relativ stark konserviert, und zum anderen muss sie auch – zumindest für CD4 – zugänglich sein. Damit ist sie eigentlich eine ideale Zielstruktur für neutralisierende Antikörper. Und tatsächlich wurden einige bNAbs identifiziert, die genau hier binden.
Grafik: Zündorf
Nachdem die Interaktion zwischen gp120 und den entsprechenden Antikörpern genauer aufgeklärt war, wurde das gp120-Derivat RSC3 entworfen (Abb. 4). Mit diesem Protein konnten sehr spezifisch B-Zellen aus peripheren Blutzellen chronisch HIV-Infizierter isoliert werden, die ihrerseits wieder hoch wirksame bNAbs bilden. So konnte RSC3 als valider Bindungspartner für diese neutralisierenden Antikörper verifiziert werden. Ob allerdings dieses Protein als Impfstoff auch wirklich in der Lage ist, die Bildung entsprechender Antikörper zu induzieren, muss sich in entsprechenden Studien erst noch zeigen.
Ein anderes gp120-Epitop für die Bildung von bNABs ist der V2/V3-Loop. Das Problem mit diesem Epitop besteht darin, dass es in der Bildung der nativen, trimeren Struktur des Env-Proteins auf der Virus-Oberfläche involviert ist. Soll also das V2/V3-Epitop als Immunogen genutzt werden, muss irgendwie die Trimerisierung von Env berücksichtigt werden. Diese Aufgabe wird allerdings dadurch erschwert, dass das native Env-Trimer sehr instabil ist und auch sein muss, da wegen der nötigen Konformationsänderungen beim Andocken an und beim Eindringen in die Wirtszelle eine beachtliche strukturelle Flexibilität notwendig ist. Darüber hinaus zeigt gerade das Env-Protein eine sehr hohe Variationsbreite zwischen den HIV-1-Subtypen, so dass es schwierig ist, einen breit wirksamen Impfstoff gegen mehrere Subtypen zu entwickeln.
Verschiedene Ansätze, stabilisierte Env-Trimere als Antigene einzusetzen, lieferten bisher keine breit wirksamen, neutralisierenden Antikörper. Und selbst die Expression des Env-Trimers auf Virus-ähnlichen Partikeln, die dann als sehr immunogener Impfstoff eingesetzt werden könnten, hat gezeigt, dass die wenigsten Moleküle nativ exponiert wurden und somit keine neutralisierende Immunantwort induziert wurde.
Grafik: Zündorf
Fazit und Ausblick
30 Jahre nach dem ersten gehäuften Auftreten von AIDS kann man resümieren: Es hat sich unglaublich viel auf dem Gebiet der Forschung an HIV und an einem Impfstoff gegen HIV getan. Gibt man in Pubmed den Suchbegriff "HIV" ein, findet man 232.130 Zitationen und unter dem Begriff "HIV vaccine" auch immerhin noch 12.966 Zitationen (Stand: 11.11. 2011)! Mit einer derartigen Publikationsleistung wird wahrscheinlich kaum ein anderes Pathogen beforscht.
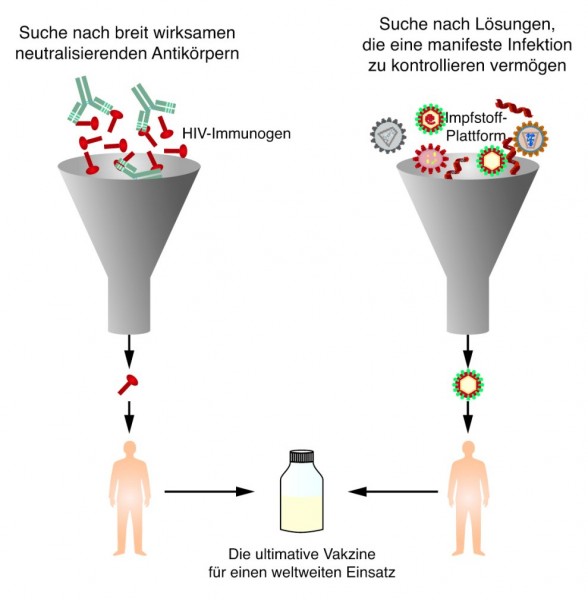
Und trotzdem ist der Erfolg bei der Impfstoffentwicklung bisher eher mäßig. Der Erkenntnisgewinn ist allerdings bereits enorm. Jedoch ist HIV ein wirklich "harter Brocken": Als übliche Eintrittspforte nutzt HIV die Schleimhäute im Anogenitalbereich. Innerhalb kurzer Zeit verschwindet das Virus in den infizierten Zellen und persistiert anschließend in einem teils sehr langlebigen Zellreservoir. Will man also wirklich die initiale Infektion verhindern, sollten möglichst in der Schleimhaut ausreichend neutralisierende Antikörper vorhanden sein, die in der Lage sind, hier bereits die Viruspartikel abzufangen. Derartige HIV-präventive Impfstoffe werden mittlerweile von sogenannten HIV-therapeutischen Impfstoffen unterschieden (Abb. 5). Mit HIV-therapeutischen Impfstoffen will man das Immunsystem von HIV-positiven Patienten in die Lage versetzen, die Infektion besser zu kontrollieren. Auch auf diesem Gebiet werden hochinteressante Ansätze verfolgt. Entsprechende Studien sowie weitere Informationen sind z. B. unter http://aidsinfo.nih.gov/vaccines/ zu finden.
Es tut sich viel auf dem Gebiet und mit den Prime-Boost-Ansätzen, den verschiedenen interessanten antigenen Strukturen von gp120 und den mittlerweile zur Verfügung stehenden Adjuvanzien besteht die Hoffnung, dass irgendwann wirklich einmal ein wirksamer Impfstoff zur Verfügung stehen könnte.
Literatur http://www.unaids.org/en UNAIDS Report on the Global AIDS Epidemic 2010 http://www.rki.de Epidemiologisches Bulletin 21/2011 http://www.iavi.org International AIDS Vaccine Initiative, u.a. mit dem Bericht AIDS Vaccine Blueprint 2008 http://www.hiv.lanl.gov/content/index HIV-Datenbanken am Los Alamos National Laboratory http://aidsinfo.nih.gov/vaccines/ Informationsseite des U.S. Department of Health and Human Services (HHS) z. B. zu klinischen Impfstoff-Studien Haynes, B.F., Moody, M.a., Liao, H.-X., et al.: B cell responses to HIV-1 infection and vaccination: pathways to preventing infection. Trends Mol Med 17 (2011), 108 – 116. Johnston, M.I., Fauci, A.S.: HIV Vaccine Development – Improving on Natural Immunity. N Engl J Med 365 (2011), 873 – 875. McElrath, M.J., Haynes, B.F.: Induction of Immunity to Human Immunodeficiency Virus Type-1 by Vaccination. Immunity 33 (2010), 542 – 554. McMichael, A.J., Borrow, P., Tomaras, G.D., et al.: The immune response during acute HIV‑1 infection: clues for vaccine development. Nat Rev Immunol 10 (2010), 11 – 23. Mikell, I., Sather, D.N., Kalams, S.A., et al.: Characteristics of the Earliest Cross-Neutralizing Antibody Response to HIV-1. PLoS Pathog 7 (2011), e1001251. Pejchal, R., Wilson, I.A.: Structure-based vaccine design in HIV: blind men and the elephant? Curr Pharm Des 16 (2010), 3744 – 3753. Robertson, D.L., Anderson, J.P., Bradac, J.A.: HIV-1 nomenclature proposal. Science 288 (2000), 55 – 56. Tebit, D.M., Nankya, I., Arts, E.J., Gao, Y.: HIV diversity, recombination and disease progression: how does fitness "fit" into the puzzle? AIDS Rev. 9 (2007), 75 – 87. Verkoczy, L., Kelsoe, G., Moody, M.A., Haynes, B.F.: Role of immune mechanisms in induction of HIV-1 broadly neutralizing antibodies. Curr Opin Immunol 23 (2011), 383 – 390. Walker, L.M., Burton, D.R.: Rational antibody-based HIV-1 vaccine design: current approaches and future directions. Curr Opin Immunol 22 (2010), 358 – 366. Wu, X., Yang, Z.-Y., Li, Y., et al.: Rational Design of Envelope Identifies Broadly Neutralizing Human Monoclonal Antibodies to HIV-1. Science 329 (2010), 856 – 861.
Autoren
Dr. Ilse Zündorf, Prof. Dr. Theodor Dingermann, Institut für Pharmazeutische Biologie, Universität Frankfurt, Max-von-Laue-Str. 9, 60438 Frankfurt/M.
Zurück zum Inhaltsverzeichnis "Schwerpunkt HIV und AIDS"



0 Kommentare
Das Kommentieren ist aktuell nicht möglich.