


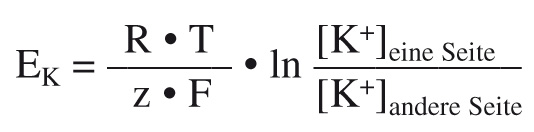
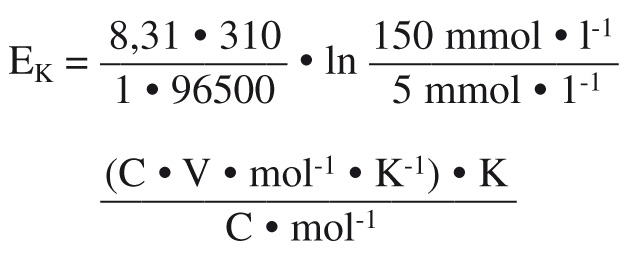













- DAZ.online
- DAZ / AZ
- DAZ 3/2013
- Die Zelle und ihr ...
UniDAZ
Die Zelle und ihr Potenzial
EditorialLiebe Leserinnen, liebe Leser! Jeder von uns besteht aus 10 bis 100 Billionen Zellen, die auf vielfältige Weise miteinander kommunizieren. So besitzt jede Zelle Rezeptoren zum Empfang von Signalen sowie Transporter zur Aufnahme und Abgabe von Stoffen; zudem befinden sich in jeder Zellmembran spannungsabhängige Kanäle für die Passage von Ionen. Während Rezeptoren und Transporter erst in den letzten Jahrzehnten identifiziert und in ihrer Struktur aufgeklärt wurden, wurde die "tierische Elektrizität" schon vor über 200 Jahren entdeckt; allerdings verstand man ihr Wesen damals noch nicht, denn Ionen waren ebenso unbekannt wie Körperzellen. Im Jahr 1887 wurde das erste EKG aufgezeichnet (sieben Jahre, nachdem Edison seine Glühlampe patentiert hatte), und nun etablierte sich die Elektrophysiologie im Grenzbereich zwischen Medizin und Physik. Heute weiß man nicht nur, wie ein Herzschlag zustande kommt und warum er sich ständig wiederholt, sondern man weiß auch, wie krankhafte Störungen dieses Rhythmus entstehen und wie entsprechende Arzneimittel (Antiarrhythmika) wirken. Lesen Sie dazu diesen Aufsatz – er ist lehrreich und spannend zugleich! Ihre UniDAZ-Redaktion |
Was hat ein Apotheker davon, das Membranpotenzial zu verstehen? Ganz allgemein kann man sagen: Das Membranpotenzial ist die Grundlage für viele physiologische Prozesse. Aber Membranpotenzial ist noch mehr: Es ist spannend! Es ist schön zu wissen, dass man mit dem Elektrokardiogramm (EKG) die Herzaktivität messen kann. Richtig toll ist es aber, wenn man weiß, warum das so ist und warum ein Medikament die QT-Zeit verlängern kann. Dass bei multipler Sklerose die Nervenleitungsgeschwindigkeit (zu) langsam ist, ist so leicht zu verstehen wie die Tatsache, dass man mit einem Liter Wasser zwar 50 Schnapsgläser füllen kann, aber nur vier Viertele-Gläser. Der Zahnarzt manipuliert das Membranpotenzial, und Sie haben keine Schmerzen bei der Behandlung. Wieso? Wie kann es sein, dass ein Ionenkanal dafür sorgt, dass eine Maus im Winter nicht friert? Wenn Sie diesen Text gelesen haben, wissen Sie es.
Abb. 1: Grundstruktur der Phospholipide. Sie bestehen aus einem Glycerolrest (rot) mit zwei daran veresterten langen Fettsäureketten, die den hydrophoben Teil bilden (gelb unterlegt). An der dritten Hydroxylgruppe des Glycerols ist eine Phosphorsäure verestert (oben), an der in der Regel noch ein Rest (R) hängt (z. B. Cholin oder Inositol). Der blau markierte Teil des Phospholipids ist hydrophil.
Keine Zelle ohne Membranpotenzial
Jede Zelle ist mit der Notwendigkeit konfrontiert, energiereiche Stoffe über die Barriere der Zellmembran aufzunehmen und Abfallstoffe abzugeben. Auf- und Abbau von Speichermolekülen verändern die intrazelluläre osmotische Konzentration und das Zellvolumen, das daher reguliert werden muss. Schließlich ist die intrazelluläre Konzentration an Proteinanionen (Enzymen!) hoch, die Cl--Konzentration muss daher gering gehalten werden. Zudem wird Information, codiert in die Frequenz von Aktionspotenzialen, über große Strecken verlustfrei transportiert. Die Frequenz von Aktionspotenzialen lässt unsere Skelettmuskeln optimal kontrahieren, die Dauer der Aktionspotenziale des Herzens bestimmt dessen geregelte Kontraktion. Wir sehen, weil Licht das Membranpotenzial der Stäbchen und Zapfen verändert. Wir hören, fühlen, schmecken, riechen mithilfe des Membranpotenzials, wir kennen die Lage unseres Körpers im Raum durch dessen Änderung und können mit seiner Hilfe die Relation unserer Hände und Füße zum Körper bestimmen. Unser Darm transportiert Nährstoffe ins Blut, die Leber entgiftet das Blut, die Nieren filtern und reinigen es, wir schwitzen, lachen, weinen, alles mithilfe des Zellmembranpotenzials. Und wir denken mit dem Membranpotenzial!
Die Funktion der Zellen und damit auch der Organe des menschlichen Körpers ist also durch die Funktion von Ionenkanälen bestimmt. Die gute extrazelluläre Zugänglichkeit der in der Zellmembran liegenden Ionenkanäle macht sie zudem nicht nur zu "drug targets" sondern auch zu guten "druggable targets".
Was ist eine Membran?
Es war einmal, vor vielen Millionen Jahren, die Ursuppe der Ozeane auf dem Planeten Erde, die bestand aus Wasser und vielen darin gelösten Teilchen. Das waren hydrophile Teilchen, z. B. Ionen oder auch organische und anorganische Moleküle. Es gab allerdings auch Teilchen, die nicht in Wasser löslich waren, die waren hydrophob.
Und dann gab es Teilchen oder besser Moleküle, die waren beides, hydrophob und hydrophil und diese nennt man amphiphil. Abbildung 1 zeigt ein solches Molekül, ein Phospholipid. Es besteht aus einem Glycerolrest (rot) mit zwei daran veresterten langen Fettsäureketten (gelb unterlegt). Die Fettsäureketten bilden den hydrophoben Teil. An der dritten Hydroxylgruppe des Glycerols ist eine Phosphorsäure verestert, an der in der Regel noch ein Rest (R) hängt (z. B. Cholin oder Inositol). Der blau markierte Teil des Phospholipids ist hydrophil.
Amphiphilen Molekülen in Wasser bleibt nun gar nichts anderes übrig, als sich zusammenzulagern, entweder als Mizellen (Abb. 2a) oder als Liposom, das ist ein flüssigkeitsgefüllter Hohlraum, der von einer Lipiddoppelschicht oder Membran umschlossen wird (Abb. 2b). So sind die hydrophoben Teile der Moleküle sozusagen "unter sich" und die hydrophilen Teile interagieren mit dem Wasser und halten das Ganze in Lösung.
Abb. 2: Kugelförmig zusammengelagerte amphiphile Moleküle
(gelb = hydrophob; blau = hydrophil), jeweils Querschnitt:
Mizelle (links) und Liposom, das mit einer Lipiddoppelschicht oder Membran einen Hohlraum umschließt (rechts).
Liposomen waren nun in der Evolution was ganz was Besonderes, denn plötzlich gab es ein "Innen", das gegen "Außen" abgegrenzt war und zwar durch eine Schicht, die für Wasser, Ionen und andere hydrophile Teilchen undurchlässig war. Innen und Außen konnten daher verschieden sein, es gab plötzlich Reaktionsräume, in denen Moleküle zusammen kommen und in einer Weise reagieren konnten, die in der Weite des Ozeans unmöglich oder sagen wir besser sehr unwahrscheinlich war. Die Reaktionsprodukte blieben dann auch noch eng beieinander und konnten weiter interagieren.
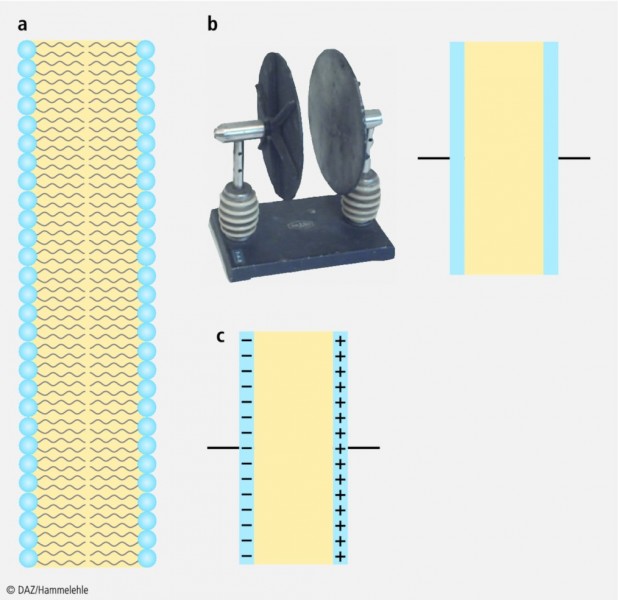
Aber das ist noch nicht einmal das Tollste an Liposomen. Nein, das Tollste an Liposomen, das ist die Membran die sie umschließt! Wenn wir uns ein Stück Membran anschauen (Abb. 3a), dann stellen wir fest, dass sie wie ein Plattenkondensator (Abb. 3b) aufgebaut ist: Je eine elektrisch geladene oder ladbare Schicht innen und außen, getrennt durch die isolierende Schicht Luft oder bei der Membran der Fettsäureketten. Das bedeutet, die Membran ist ein Kondensator. Was genau aber ist ein Kondensator, und wo bleibt das Membranpotenzial?
Abb. 3:
Abschnitt einer Lipiddoppelschicht oder Membran (links)
sowie ein Plattenkondensator im ungeladenen Zustand (rechts)
und im geladenen Zustand (unten).
FrageWas war zuerst da, die Zelle oder ihr Membranpotenzial? Das Membranpotenzial, denn auch Liposomen können ein Membranpotenzial haben, weil ihre Membran ein Kondensator ist. An einem Liposom wird jedoch ein Strom nur zufällig den Kondensator laden, während dies an einer Zelle durch gerichtete Prozesse geschieht. |
Was ist ein Kondensator?
Nichts anderes als ein Ladungsspeicher. Die Membran speichert die elektrische Ladung, die das Membranpotenzial ausmacht.
Wenn wir uns z. B. eine wässrige Salzlösung anschauen, dann ist die Anzahl der Kationen (positiv geladene Teilchen) genauso groß wie die Anzahl der Anionen (negativ geladen). Eine Ladungstrennung ist nicht möglich, d. h., es gibt keine Lösung, in der eine Sorte Ionen überwiegt, die also per se geladen wäre.
Bei einem Kondensator ist das anders, hier ist durchaus eine Ladungstrennung möglich (Abb. 3c). Wenn man eine Seite des Kondensators negativ lädt, dann ist die andere Seite im Vergleich dazu positiv geladen. In diesem Zustand befinden sich beide Seiten auf unterschiedlichem Potenzial, wir haben also eine Potenzialdifferenz E (meistens lässt man "differenz" aus Bequemlichkeit weg und sagt nur "Potenzial") oder auch elektrische Spannung U. Wie viel Ladung Q man für eine bestimmte Spannung U braucht, hängt von der "Größe" des Kondensators ab, man spricht von seiner Kapazität C. Es gilt:
Q = C • U
Das heißt, je größer die Kapazität C des Kondensators ist, desto mehr Ladung Q muss ich in ihn "einfüllen", um eine bestimmte Spannung U zu erreichen.
Für einen Plattenkondensator, wie es die Membran eines Liposoms bzw. einer Zelle ist, errechnet sich die Kapazität in erster Näherung auch so:
C = A/d
Dabei ist A die Fläche der Kondensatorplatten, also die Größe der Zelle (bzw. ihrer "Hülle", der Membran) und d (im Nenner!) der Abstand der Platten, also die Dicke der Membran. Dies zu wissen, wird dann wichtig, wenn wir uns überlegen, warum Myelinscheiden an Nervenfasern die Leitungsgeschwindigkeit erhöhen. Dann erhöht nämlich die Myelinschicht, die auf der Membran aufliegt, den Plattenabstand d, und der Kondensator hat eine viel geringere Kapazität.
Wie lädt man einen Kondensator?
Ganz einfach, man muss Ladung in ihn "einfüllen". Im Prinzip hat das jeder schon mal gemacht, zumindest alle, die den Akku z. B. ihres Handys schon mal geladen haben. Auch in einem Akku (genauer: Akkumulator (= Anhäufer, Sammler)) gibt es zwei Kompartimente, die Ladung aufnehmen können. Diese sind durch eine Isolierschicht voneinander getrennt, also ganz ähnlich wie bei einem Kondensator, wie wir ihn hier betrachten. Zusätzlich benötigt man eine Spannungsquelle, also das Ladegerät, ein doppeltes Ladekabel, und auf geht’s. Aber wie?
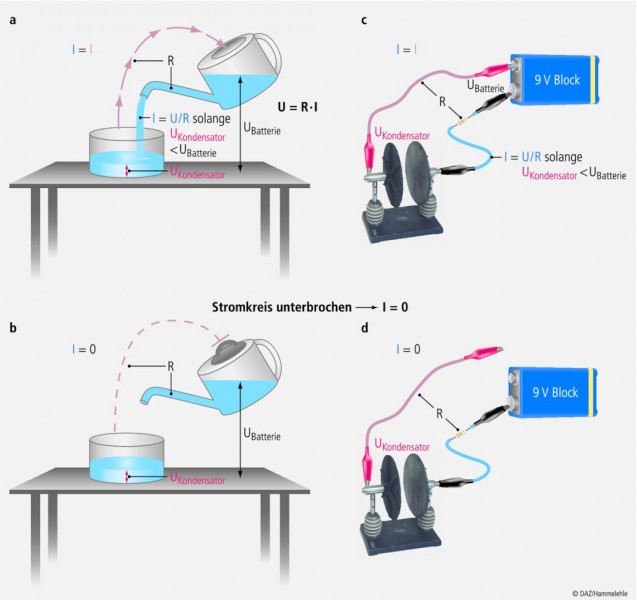
Schauen wir uns zunächst an, was passiert, wenn wir aus einer Kanne Wasser in ein Glas gießen wollen (Abb. 4a): Als Erstes müssen wir die Kanne anheben! Wir brauchen eine Potenzialdifferenz zwischen dem Wasser in der Kanne und dem Glas. In Analogie zum elektrischen Stromkreis habe ich diese Potenzialdifferenz UBatterie genannt (Abb. 4a). Wenn wir durch diese Potenzialdifferenz nun eine Triebkraft haben, können wir einen Wasserstrom I in das Glas strömen lassen, vorausgesetzt, es gibt einen Weg dafür, eine Leitfähigkeit G. Die Leitfähigkeit hat immer auch eine bestimmte Größe: Ist die Tülle der Kanne weit, so fließt viel Strom, weil ihr Widerstand klein ist, und umgekehrt. Eine Leitfähigkeit ist also immer auch ein Widerstand R, beide Größen sind umgekehrt proportional: R = 1/G.
In Abb. 4a ist dies die Tülle der Kanne und der Raum bis zur Oberfläche des Wassers im Glas. Je mehr sich das Glas füllt, desto größer wird auch die Potenzialdifferenz zwischen der Oberfläche des Wassers und dem Boden des Glases, die ich in Abb. 4a UKondensator getauft habe. Es fließt so lange (Wasser-)Strom (I), wie UKondensator kleiner als UBatterie ist.
Die rosa Pfeile in Abb. 4a zeigen, dass es aber auch einen zweiten Widerstand geben muss, damit der Wasserstrom fließen kann. Nur wenn die Luft aus dem Glas in die Kanne gelangen kann, der Stromkreis also geschlossen ist, kann Wasserstrom fließen! Wird der Stromkreis an irgendeiner beliebigen Stelle unterbrochen (Abb. 4b), dann kann kein Strom mehr fließen. Probieren Sie es aus, verschließen Sie nur die Einfüllöffnung Ihrer Gießkanne luftdicht und Ihre Blumen und Pflanzen vertrocknen.
Abb. 4: Stromfluss
von der Gießkanne in den Topf (Wasser) bzw. von der Batterie zum Kondensator. Der Luftstrom bzw. ein zweites Kabel schließt den Stromkreis (oben). U = Spannung (Potenzial); R = Widerstand; I = Stromfluss.
So wie man nun Wasser in ein Glas gießen kann, so kann man in einen Kondensator elektrische Ladung füllen. Wir brauchen dazu eine Gleichspannungsquelle, also z. B. eine Batterie, die uns die Triebkraft und die Ladung liefert. Dann brauchen wir zwei Widerstände, z. B. Kabel, um einen geschlossenen Stromkreis herzustellen, und schließlich den Kondensator, in den man die Ladung füllen kann (Abb. 4c). Es wird zunächst viel Strom fließen, und der ist überall im Stromkreis gleich stark (Iblau = Irosa). Der Strom wird aber exponenziell abnehmen, und zwar in dem Maß, wie UKondensator zunimmt. Ist schließlich UKondensator gleich groß wie UBatterie, fließt kein Strom mehr, wir haben ein Gleichgewicht zwischen der Triebkraft der Batterie und der Spannung des Kondensators erreicht. Bei einer Zelle nennt man dies das Gleichgewichtspotenzial. Man spricht auch von Nullstrompotenzial, eben weil kein Strom mehr fließt, oder von Umkehrpotenzial, weil oberhalb dieses Potenzials der Strom in die eine Richtung fließt, unterhalb in die andere. Ein viertes Synonym ist das Nernstpotenzial, zu dem wir im nächsten Kapitel kommen.
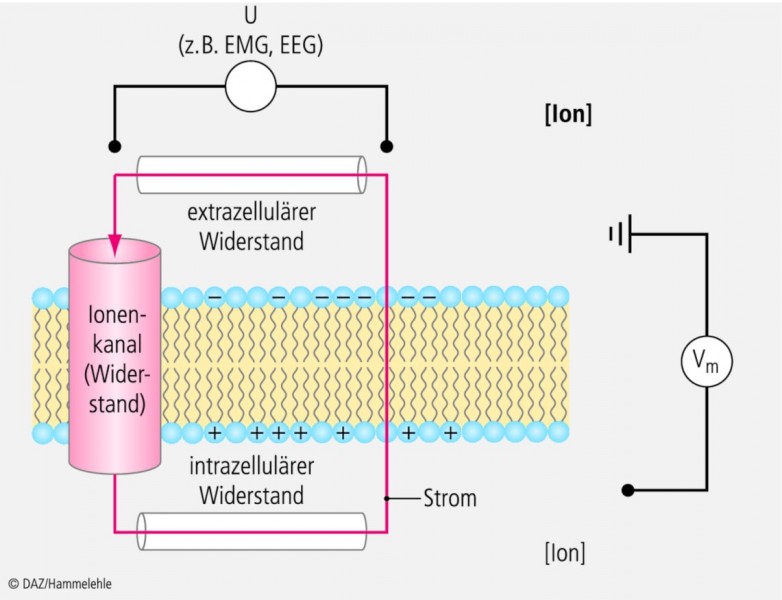
Bleibt noch zu erwähnen, dass auch hier kein Strom fließt, wenn der Stromkreis irgendwo unterbrochen wird (Abb. 4b). Und das ist der Clou an der Sache! Das heißt nämlich, wenn Strom über einen Ionenkanal in der Membran fließt (Abb. 5), der die Spannung des Membrankondensators (also das Membranpotenzial Vm) ändert, dann muss unbedingt auch Strom im Extrazellularraum und im Intrazellularraum fließen, anders geht es nicht, wir brauchen einen geschlossenen Stromkreis! Und das heißt: Wenn viele Zellen gleichzeitig elektrische Aktivität zeigen, z. B. Herzmuskelzellen oder Neuronen im Gehirn, dann fließt so viel Strom durch den extrazellulären Widerstand, dass die Spannung die er erzeugt (U = R • I) mit Elektroden auf der Haut gemessen werden kann. Zeichnet man diese Spannung mit einem Schreiber auf, erhält man ein Elektrokardiogramm (EKG), ein Elektroenzephalogramm (EEG) oder auch ein Elektromyogramm (EMG, z. B. von einem Armmuskel). Wenn Sie das verstanden haben, dann können Sie jetzt leicht herausfinden, wann man im EEG große langsame alpha-Wellen misst:
A) Im Wachsein, wenn viele kleine Gehirnareale unterschiedliche Aufgaben wahrnehmen;
B) im Schlaf, wenn große Gehirnareale gleiche Aktivität zeigen.
Wo aber kommen die Batterien her, die den Membrankondensator einer Zelle laden?
Abb. 5:
Wenn Strom durch einen Ionenkanal in der Zellmembran fließt, fließt er auch durch den extra- und intrazellulären Widerstand und erzeugt dort ein Potenzial. Die Spannung (U), die am extrazellulären Widerstand erzeugt wird, kann man als EKG, EEG oder EMG messen.
Das Nernstpotenzial
Als sich aus Liposomen die ersten Zellen zu formen begannen, hatten sie das gemeinsame Problem der "osmotischen Krisen" [1]. Durch erste Stoffwechselprozesse oder durch zufälligen Einstrom von Ionen und/oder Wasser entstand die Notwendigkeit, Ionen (v. a. H+, aber auch Na+ oder K+) aktiv über Membranen transportieren zu müssen, um zu starken Anreicherungen entgegenzuwirken. Ionenpumpen (ATPasen) zählen daher zu den ältesten Enzymen überhaupt. Diese befördern unter ATP-Verbrauch Ionen auch entgegen Konzentrationsgradienten über Membranen. Aber auch das Gegenteil ist realisiert, die F1/F0-ATPase nutzt den H+-Gradienten über die innere Mitochondrienmembran aus, um aus ADP und Pi ATP zu generieren.
Für das Verständnis des Membranpotenzials ist die Na+ /K+-ATPase von Bedeutung. Diese reichert K+ in den Zellen an, das aus dem Außenraum in die Zellen gepumpt wird, und pumpt gleichzeitig Na+ aus den Zellen heraus.
Die Konzentrationsgradienten für Na+ und K+ stellen also die chemischen Triebkräfte für die Ströme dieser Ionen, sie müssen daher die "Batterien" (Na+-Batterie, K+-Batterie) sein, die den Membrankondensator laden. Wenn das so ist, dann muss es eine Möglichkeit geben, Ionenkonzentrationsgradienten in elektrische Spannung umzurechnen. Und hier kommt jetzt Professor Nernst ins Spiel.
Walther Nernst (1864 – 1941) war Physiker und Chemiker, der für seine Arbeiten in der Thermochemie 1920 den Nobelpreis erhalten hat. Seine Überlegungen zur Konzentrationsabhängigkeit der Spannung in einer galvanischen Zelle führten zur Nernst-Gleichung, deren Sonderform es uns ermöglicht, die Spannung eines Ionenkonzentrationsgradienten zu errechnen. Die Formel für z. B. K+ lautet:
Bitte jetzt nicht aufgeben, die Formel scheint nur schwierig zu sein, in Wirklichkeit ist sie ganz einfach! R ist die universelle Gaskonstante (8,31 C • V • mol-1 • K-1); T ist die Temperatur in Kelvin (K; bei einer Säugerzelle mit 37 °C also ~310 K); z die Wertigkeit des Ions (da K+ einfach positiv ist also 1); F die Faradaykonstante (96.500 C • mol-1) und [K+] die Konzentration von K+ auf der jeweiligen Seite der Membran.
Wenn man dies einfach mal hinschreibt, gleich nach Werten und Dimensionen getrennt, kriegt man:
Durch Kürzen wird daraus:
EK = 0,027 • ln 30 V
Die Einheit ist also in der Tat Volt, d. h. wir können die Spannung der "Batterie" ausrechnen, die von einem Ionenkonzentrationsgradienten gebildet wird! Da Professor Nernst keinen Taschenrechner besaß, wohl aber dekadische Logarithmentafeln, wurde die Formel noch ein wenig umgerechnet, bleibt aber einfach. Zudem wird das Membranpotenzial immer in mV angegeben. Es ergibt sich also rund gerechnet und absolut gesehen (also unabhängig vom Vorzeichen, deshalb die senkrechten Striche):
|EK| ~60 • lg 30 mV oder
|EK| ~60 • 1,5 mV = 90 mV
K+ strömt von innen nach außen, d. h. der Membrankondensator wird eigentlich außen positiv geladen. In der Physiologie ist nun das Potenzial des Extrazellularraums per Definition 0 mV, weil die Badelektrode auf das Erdpotenzial gelegt wird. Was außen also nicht positiv sein kann, ist daher innen negativ! Deshalb ist:
EK = ‑90 mV
Wenn man sich also die vereinfachte Nernst-Gleichung (für einwertige Ionen)
EX = 60 • lg [X]/[X] (mV)
gut merkt, kann man sehr schnell und einfach Gleichgewichtspotenziale für Ionen ausrechnen, z. B.
ENa = 60 • lg 140/14 mV = 60 mV
ZusammenfassungDie Lipiddoppelschicht der Membran ist physikalisch gesehen ein Kondensator. Ionenkonzentrationsgradienten entsprechen Batterien, mit deren Hilfe dieser Kondensator durch einen Stromfluss aufgeladen wird. Dabei fließt der Ionenstrom durch entsprechende Ionenkanäle in der Membran und im Intra- und Extrazellularraum, also in einem geschlossenen Stromkreis (Abb. 4 a), und zwar so lange, bis das Nullstrompotenzial erreicht ist, die Membran also das gleiche Potenzial hat wie die Gradientenbatterie. Dies gilt, wenn es nur für eine Ionensorte selektive Kanäle gibt, und zwar unabhängig von der Anzahl der Kanäle (wird die Tülle der Kanne in Abb. 4 a enger, dann dauert es zwar länger, bis der Topf voll ist, aber man kann gleich viel Wasser einfüllen). |
Wie viel Strom fließt denn nun?
Das Ohmsche Gesetz stellt einen sehr einfachen Zusammenhang zwischen Spannung U, Widerstand R und Strom I her:
U = R • I (oder umgeformt I = U/R oder I = U • G)
Wenn wir das auf die Membran für ein Ion übertragen (und statt Widerstand die Leitfähigkeit (G = 1/R) nehmen), dann ist die Spannung, die das Ion spürt das aktuelle Membranpotenzial Vm minus Nernstpotenzial des Ions EIon. Es ergibt sich daraus (auch wenn‘s komisch aussieht, das ist immer noch das Ohmsche Gesetz):
IIon = (Vm – EIon) • GIon
Wenn Vm = EIon ist, dann ist IIon = 0 (Nernstpotenzial), wenn aber Vm ≠ EIon, dann spielt, wie wir sehen, die Leitfähigkeit GIon eine Rolle.
Was passiert, wenn zwei Ionensorten strömen?
Welches Vm stellt sich ein, wenn zwei Ionen beteiligt sind, also z. B. Na+ und K+? Generell gilt, dass die Summe aller Ströme bei jedem beliebigen Vm gleich Null ist (wäre es nicht so, würde der Strom die Spannung ändern!). Also ist in unserem Beispiel
INa + IK = 0 oder INa = IK.
Betrachten wir den einfachsten Fall, nämlich dass die beiden Leitfähigkeiten gleich sind, dann muss die Triebkraft für beide Ionen (Vm – EIon) auch gleich sein. Profis können das nun per Gleichung ausrechnen, wir anderen schauen auf Abbildung 6a.
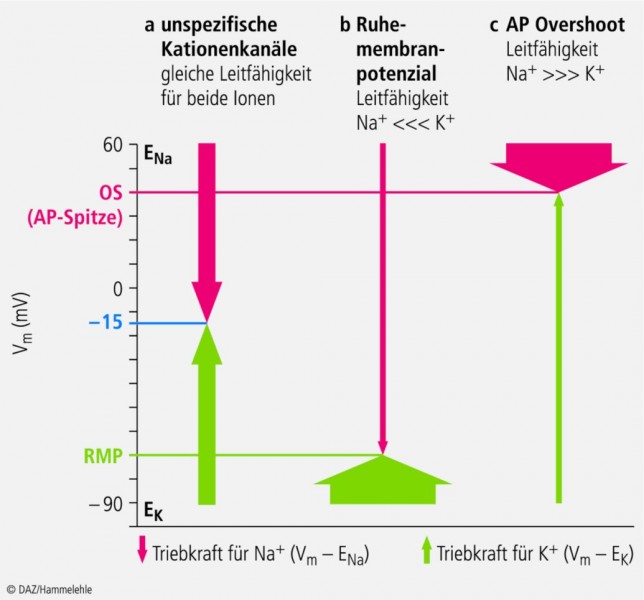
Abb. 6:
Wenn die Leitfähigkeit für Na+
und K+
gleich groß ist (die Pfeile sind gleich dick), dann ist Vm
= -15 mV, also gleich weit von EK
und ENa
entfernt (a). Am Ruhemembranpotenzial ist die Leitfähigkeit für K+
groß (dicker Pfeil) bei kleiner Triebkraft, die für Na+
klein (dünner Pfeil) bei großer Triebkraft (b).
An der AP-Spitze (Overshoot) ist das Verhältnis gerade umgekehrt (c).
Wenn also die Leitfähigkeit für Na+ und K+ gleich groß ist (die Pfeile sind gleich dick), dann ist Vm = ‑15 mV, also gleich weit von EK und ENa entfernt. Gibt es denn so etwas? Ja, es gibt unspezifische Kationenkanäle, die für Na+ und K+ in etwa gleich gut leitend sind. Wenn diese geöffnet werden, versuchen sie Vm nach ‑15 mV zu bringen. Dies gilt für Liganden-gesteuerte Kanäle wie z. B. den nicotinischen Acetylcholinrezeptor (Skelettmuskeln) oder für HCN-Kanäle, die durch Hyperpolarisation geöffnet werden (z. B. im Sinusknoten des Herzens). Es ist in der Regel die Öffnung solcher unspezifischer Kationenkanäle, die dadurch zur Erregung von Zellen führt, dass Vm vom Ruhemembranpotenzial RMP zum Schwellenpotenzial SP depolarisiert wird (siehe unten).
Das Ruhemembranpotenzial erregbarer Zellen liegt in etwa zwischen ‑70 mV und ‑80 mV. Da die Summe beider Ströme (Na+ und K+) auch hier gleich sein muss, ist also in Ruhe die K+-Leitfähigkeit groß (dicker Pfeil bei kleiner Triebkraft) und die für Na+ sehr klein (dünner Pfeil bei großer Triebkraft) (Abb. 6b).
Zusammenfassend können wir uns das ganz einfach vorstellen, wenn wir einen Mischwasserhahn betrachten. Dreht man nur den Kaltwasserhahn oder nur den Heißwasserhahn auf, ist die Temperatur unabhängig von der Stärke des Wasserstroms, wir sind also bei "EK " oder "ENa". Werden beide Hähne zugleich aufgedreht, dann ist die Mischtemperatur abhängig von den einzelnen Temperaturen (heiß und kalt) und vom Anteil der einzelnen Wasserströme am Gesamtstrom. Um im Bild zu bleiben: Werden beide Hähne gleich weit aufgedreht, sind wir bei ‑15 mV (Abb. 6a).
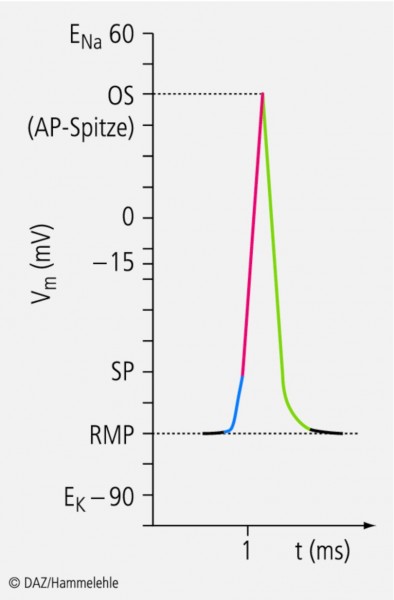
Abb. 7: Depolarisation und Repolarisation einer Zelle
Durch das Öffnen unspezifischer Kationenkanäle steigt das Membranpotenzial Vm
vom Ruhemembranpotenzial (RMP) zum Schwellenpotenzial (SP; blaue Linie); dann strömt durch Na+-Kanäle Na+
in die Zelle ein, und das Potenzial steigt steil bis zum Overshoot an (OS; rote Linie). Danach strömt durch K+-Kanäle
K+ aus der Zelle, und die Zelle erreicht wieder ihr Ruhemembranpotenzial (grüne Linie).
Erregbare Zellen und das Aktionspotenzial
Das bisher Gesagte gilt für alle Zellen, da alle Zellen Ionenkanäle und ein Membranpotenzial für ihre unterschiedlichen Aufgaben haben. Ab jetzt werden wir aber unterscheiden zwischen erregbaren Zellen (z. B. Muskelfasern, Neuronen) und nicht-erregbaren Zellen (z. B. Leberzellen, Nierentubuluszellen).
Erregbare Zellen zeichnen sich dadurch aus, dass sie Aktionspotenziale bilden können. Sie brauchen dazu Kanäle die spannungsabhängig sind, das heißt, dass sie sich auf eine Veränderung des Membranpotenzials öffnen. Für das generelle Verständnis der Mechanismen, die Aktionspotenzialen zugrunde liegen, brauchen wir zunächst drei Typen von spannungsabhängigen Kanälen, Na+-, K+- und Ca2+-Kanäle. Diese spannungsabhängigen Kanäle öffnen auf eine Depolarisation vom RMP zum Schwellenpotenzial (SP; ‑55 bis ‑40 mV, abhängig von Zelle und Kanal).
Wie alle Aktionspotenziale (APs) wird auch das Na+-Aktionspotenzial durch eine Depolarisation zum SP ausgelöst. Die Depolarisation erfolgt wiederum durch das Öffnen unspezifischer Kationenkanäle (blauer Teil des APs, Abb. 7a), die zumeist durch Liganden, z. B. Neurotransmitter, aktiviert werden und ihr Nullstrompotenzial bei ‑15 mV haben (siehe oben und Abb. 6a). Bevor jedoch dieses Potenzial erreicht wird, wird das SP überschritten, und es öffnen spezifische, spannungsabhängige Na+-Kanäle. Durch den Na+-Einstrom wird jetzt auf die oben beschriebene Weise der Membrankondensator auf der Innenseite positiv geladen (roter Teil des APs, Abb. 7a). Es kommt zu einer Potenzialumkehr, und Vm erreicht maximale Werte von ca. +40 mV (sog. Overshootpotenzial, Abb. 6c und 7a).
Das Besondere an spannungsabhängigen Na+-Kanälen ist nun, dass der Offenzustand nur von kurzer Dauer ist und unmittelbar darauf ein neuer Geschlossenzustand eingenommen wird. In diesem neuen Geschlossenzustand nennt man die Kanäle inaktiviert oder refraktär. Die Na+-Kanäle verbleiben in diesem Zustand, solange die Membran depolarisiert ist. Allerdings öffnen beim Überschreiten des SPs nicht nur die Na+, sondern auch spannungsabhängige K+-Kanäle, die die Membran wieder repolarisieren. Diese Öffnung ist jedoch etwas verzögert, sodass der Na+-Einstrom zunächst nicht oder nur unwesentlich behindert wird. Erst wenn die Na+-Kanäle inaktiviert sind, kommt der K+-Strom zum Tragen, und der Membrankondensator wird wieder negativ umgeladen (grüner Teil des APs, Abb. 7a). Der Strom fließt also bei Beginn der Erregung, wenn die Na+-Kanäle öffnen, in die eine Richtung und danach aufgrund des K+-Stroms entgegengesetzt.
Wenn der Membrankondensator wieder auf Werte nahe des RMPs geladen ist, beginnt die Erholung der Na+-Kanäle von der Refraktärzeit. In dieser Phase gehen die Na+-Kanäle direkt vom geschlossen-inaktivierten in den geschlossen-aktivierbaren Zustand über. Danach können sie durch eine erneute Depolarisation geöffnet werden. Solange alle Na+-Kanäle inaktiviert sind spricht man von absoluter Refraktärzeit. Können einige Kanäle schon wieder öffnen, andere noch nicht, dann heißt das relative Refraktärzeit. Die Refraktärzeit ist vorüber, wenn alle bei diesem Membranpotenzial aktivierbaren Kanäle wieder öffnen können.
Das Na+-Aktionspotenzial breitet sich aus
Wenn wir uns eine Kugel voller spannungsabhängiger Na+- und K+-Kanäle vorstellen, dann wird ein AP, das am Nordpol entsteht (Abb. 8a), sich nach allen Richtungen über die Kugel ausbreiten und am Südpol erlöschen.
Abb. 8:
Ein Aktionspotenzial, das am Nordpol entsteht (links; blau) breitet sich nach Süden aus (rote Pfeile).
In einer Membran breitet sich das AP gezielt in die Richtung aus, wo die Ionenkanäle aktivierbar sind (rechts).
Warum ist das so? Nun wir wissen, dass der Strom, der durch einen Ionenkanal fließt, immer in einem geschlossenen Stromkreis fließt (Abb. 5). Der Einstrom von Na+ während des Nordpol-APs sorgt also dafür, dass der Membrankondensator in der Umgebung des Pols positiv geladen wird. Diese Depolarisation öffnet erneut Na+-Kanäle, es kommt wieder zum Einstrom von Na+, was wieder zur Depolarisation der Umgebung der Stelle führt, an der gerade Na+-Einstrom stattfindet.
Hemmt nun der Zahnarzt mit einem Lokalanästhetikum die Na+-Kanäle des Zahnnerven, dann werden die Schmerzrezeptoren am Zahn noch reagieren, aber ihre Signale, in Form von APs, werden nicht mehr zum Gehirn weitergeleitet; man spürt also keinen Schmerz.
Warum aber breitet sich das AP nur vorwärts aus, obwohl der Strom doch auch die Region hinter dem AP depolarisiert (der Strom breitet sich ja in alle Richtungen aus)? Dies zeigt Abbildung 8b: In der Richtung aus der das AP kam sind die Na+-Kanäle inaktiviert und können durch die Depolarisation nicht geöffnet werden. Ein neues AP kann also nur in Erregungsausbreitungsrichtung entstehen, in der die Na+-Kanäle aktivierbar sind. Die Refraktärzeit der Na+-Kanäle sorgt also dafür, dass sich die Erregung nur gerichtet ausbreitet.
Gerichtete Erregungsleitung im Herzen
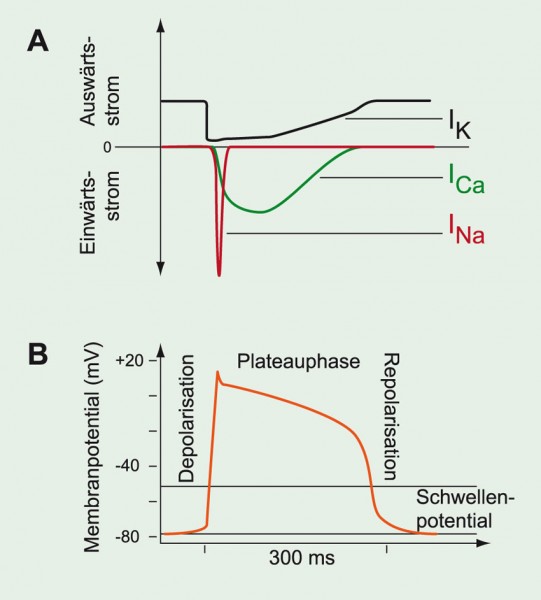
Diese Tatsache ist besonders am Herzen wichtig, bei dem das Erregungsleitungssystem (His-Bündel, Kammerschenkel, Purkinje-Fasern) das AP schnell und gerichtet zur Herzspitze leiten muss, bevor dann die Erregung über das Kammermyokard wieder zur Herzbasis läuft. Auf diese Weise kontrahieren die Ventrikel von unten nach oben und werfen so das Blut nach oben durch Pulmonal- und Aortenklappe aus. Da am Ventrikel auch spannungsabhängige Ca2+-Kanäle am AP beteiligt sind, ist das AP viel länger (~200 – 300 ms) als ein reines Na+-AP, wie man es z. B. in Skelettmuskeln findet (Abb. 9). Während der sogenannten Plateauphase des APs der Ventrikel sind die Na+-Kanäle jedoch inaktiviert, weil die Erholung von der Inaktivierung erst nach der Repolarisation der Membran beginnt (siehe oben).
Abb. 9A: Membranströme
während eines APs der Ventrikelmuskelzellen. B: Sobald das SP erreicht ist, öffnen spannungsabhängige Na+-Kanäle und sorgen für die initiale Depolarisation, die auch die Ca2+-Kanäle öffnet. Während der Plateauphase strömt Ca2+ in die Zelle, die Na+-Kanäle sind während dieser gesamten Phase inaktiviert. Beim Überschreiten des SP öffnen auch K+-Kanäle, der Strom durch diese Kanäle wird aber nur langsam größer. Wenn die K+-Leitfähigkeit größer wird als die Ca2+-Leitfähigkeit, repolarisiert die Zelle. Die spannungsabhängigen Kanäle schließen; die Na+-Kanäle beenden die Refraktärzeit und werden wieder aktivierbar.
Aus Leistner/Breckle, Pharmazeutische Biologie, 7. Aufl. 2008, S. 537.
Aus Leistner/Breckle, Pharmazeutische Biologie, 7. Aufl. 2008, S. 537.
Wie bei jedem anderen AP wird auch am Herzen die Repolarisation zum RMP durch das Öffnen spannungsabhängiger K+-Kanäle bewirkt. Hemmt nun ein Pharmakon solch einen K+-Kanaltypen (z. B. den hERG-Kanal), dann wird das AP länger, noch länger, als es ohnehin schon ist. Dies bedeutet, dass im EKG die QT-Zeit verlängert ist: Ist die Erregung am Ventrikel angekommen, so sehen wir im EKG die Q-Zacke, die den Beginn der Erregung des His-Bündels anzeigt. Die T-Welle zeigt im EKG die Repolarisation an. Da durch das Pharmakon ein Teil der repolarisierenden K+-Kanäle gehemmt ist, kommt die T-Welle aber später als gewöhnlich, die QT-Zeit ist verlängert.
Dies ist deshalb besonders gefährlich, weil die natürliche Abfolge von Erregung und Refraktärzeit während der Erregungsleitung am Ventrikel gestört sein kann. Ein verlängertes AP einer Ventrikelzelle in der Nähe der Herzbasis könnte das His-Bündel vorzeitig erregen, es käme zu einem vorzeitigen Wiedereintritt (reentry) der Erregung, mit der Folge, dass "kreisende Erregungen" ausgelöst werden können, die ein gerichtetes Pumpen des Blutes unmöglich machen. Die Kammer flimmert, das Herz steht still, und nur der Defibrillator kann noch helfen!
AP-Leitungsgeschwindigkeit
Ein Na+-AP breitet sich über z. B. eine Skelettmuskelfaser mit einer Geschwindigkeit von ~10 m/s aus. Dies scheint die maximale Leitungsgeschwindigkeit für Strukturen ohne Myelinschicht (s. u.) zu sein, da selbst im etwa 1 mm dicken Riesenaxon des Tintenfisches, der wie alle Protostomia (z. B. Schnecken, Würmer, Insekten, Spinnen) keine Myelinscheiden besitzt, die Leitungsgeschwindigkeit nicht höher ist. Da es damit bei einem 30 m langen Brontosaurus 3 s gedauert hätte, bis ein Angriff auf die Schwanzspitze im Gehirn bemerkt worden wäre, musste sich evolutionär also ein anderes Prinzip durchsetzen, um die Leitungsgeschwindigkeit zu erhöhen, als den intrazellulären Widerstand (siehe Abb. 5) durch Vergrößerung des Querschnitts zu senken.
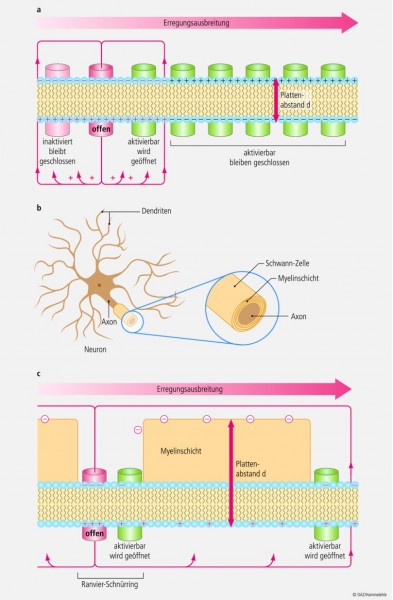
Abb. 10: Ausbreitung eines Aktionspotenzials an einem Axon;
a) an einem Axon ohne Myelinscheide wird nur jeweils der nächste aktivierbare Ionenkanal depolarisiert;
b) Nervenzelle (Neuron), deren Axon von Schwann-Zellen umgeben ist (Myelinschicht);
c) bei einem Axon mit Myelinschicht kann ein Aktionspotenzial mehrere Schnürringe in größeren Entfernungen bis zur Schwelle depolarisieren.
a) an einem Axon ohne Myelinscheide wird nur jeweils der nächste aktivierbare Ionenkanal depolarisiert;
b) Nervenzelle (Neuron), deren Axon von Schwann-Zellen umgeben ist (Myelinschicht);
c) bei einem Axon mit Myelinschicht kann ein Aktionspotenzial mehrere Schnürringe in größeren Entfernungen bis zur Schwelle depolarisieren.
Wenn man Abbildung 10a anschaut, dann fällt auf, dass die Ladungsmenge Q, die durch den offenen Kanal eingeströmt ist, gerade ausreicht, die unmittelbare Umgebung bis zum Schwellenpotenzial umzuladen, sodass gerade nur benachbarte, aktivierbare Na+-Kanäle geöffnet werden können. Erst wenn der Nachbarkanal aktiviert ist, kann wieder der nächste Kanal geöffnet werden, sodass das AP von Membranabschnitt zu Membranabschnitt sich entlang einer unmyelinisierten Axonmembran ausbreitet.
Bei Wirbeltieren allerdings sind die Axone schnell leitender Neuronen von einer Myelinscheide umgeben. Die Myelinscheide besteht aus einer langen Membranausstülpung spezieller Zellen (Schwann-Zellen, Oligodendrozyten), die in vielen Lagen eng um die Axone der Neuronen gewickelt ist (Abb. 10b). An bestimmten Stellen (alle 1 – 3 mm), den sogenannten Ranvier-Schnürringen, fehlt die Myelinschicht. Nur in der Membran der Schnürringe gibt es Kanäle. Das Ganze hat nun für die Leitungsgeschwindigkeit zwei entscheidende Vorteile:
1. Da es unter der Myelinschicht keine Kanäle gibt, sind auch keine K+-Kanäle vorhanden, durch die positive Ladung aus der Zelle fließen kann. Die Ladungsmenge an positiven Ladungen (Na+), die beim AP am Schnürring einströmt, kann somit voll für die Depolarisation der Umgebung unter der Myelinschicht genutzt werden.
2. Durch die Myelinschicht wird der Plattenabstand d des Membrankondensators sehr viel größer. Dies bedeutet, dass die Kapazität des Kondensators sehr gering ist (siehe oben), und dies wiederum heißt, dass nun sehr viel weniger Ladung gebraucht wird, um die Membran bis zum Schwellenpotenzial zu depolarisieren (Abb. 10c). Es wird also nicht nur die direkte Nachbarschaft des APs ausreichend für ein neues AP depolarisiert, sondern sehr weite Bereiche, genauer 3 – 4 Schnürringe gleichzeitig. Das AP wird also nicht von Membranabschnitt zu Membranabschnitt weitergeleitet, sondern "springt" sozusagen über 3 – 4 Schnürringe gleichzeitig weiter (ca. 1 cm!), weshalb man diesen Vorgang auch Saltatorische Erregungsleitung nennt. Und so wie ein Aufzug, der in jeder Etage hält, langsamer ist als einer, der durchfährt, so wird auch ein AP an einem myelinisierten Axon sehr viel schneller geleitet (bis zu 150 m/s).
Multiple Sklerose ist eine Autoimmunerkrankung, bei der sich das körpereigene Immunsystem gegen die Myelinscheiden der Neuronen richtet und sie abbaut. Die Myelinscheiden sind daher dünner als normal, der Plattenabstand ist geringer, d. h. dass der Membrankondensator eine größere Kapazität hat (siehe oben). Das bedeutet aber, dass jetzt in jeden Membranabschnitt sehr viel mehr Ladung "eingefüllt" werden muss, um ihn bis zum Schwellenpotenzial zu depolarisieren. Die Ladungsmenge, die während des APs an einem Schnürring einströmt, reicht also nicht so weit wie im gesunden Zustand, um wieder APs auszulösen. Es werden nicht 3 – 4 Schnürringe gleichzeitig aktiviert, sondern vielleicht gerade nur der nächste, der dann durch sein AP gerade wieder den nächsten Schnürring aktiviert. Daher ist die Leitungsgeschwindigkeit von nur noch teil-myelinisierten Axonen bei multipler Sklerose verringert.
Bleibt noch zu klären, warum die Myelinscheide Myelinscheide heißt: Rudolf Virchow (1821 – 1902) entdeckte an Nervenfasern die Markscheide und schlug vor, sie Myelin (griech. myelòs = Mark) zu nennen.
Ströme und Membranpotenzial in nicht-erregbaren Zellen
Wie alle Zellen sind natürlich auch nicht-erregbare Zellen auf das Membranpotenzial angewiesen. An diesen Zellen dient das Membranpotenzial aber nicht direkt der Signalverarbeitung, sondern vorwiegend dem (Ionen‑)Transport.
Oben haben wir schon davon gesprochen, dass das Membranpotenzial geeignet ist "osmotischen Krisen" zu begegnen und dafür brauchen es alle Zellen auch heute noch. Selbst wenn eine Körperzelle in einem osmotisch einigermaßen konstanten extrazellulären Milieu lebt, so muss sie doch aus einzelnen Bausteinen, z. B. Glucose, Glykogen aufbauen und umgekehrt. Dabei verändert sich die intrazelluläre Osmolarität, und das durch Wasserein- bzw. Wasserausstrom veränderte Zellvolumen muss durch Ionentransport über die Membran ausgeglichen werden. Schwellen die Zellen, werden v. a. K+- und Cl--Kanäle aktiviert. Wird die Leitfähigkeit und damit der Strom für K+ größer, so hyperpolarisiert die Zelle, was die Triebkraft für den Cl--Ausstrom erhöht. Schließlich folgt Wasser dem Salz aus der Zelle, zumeist über spezielle Wasserkanäle, die Aquaporine. Beim Schrumpfen von Zellen werden spezielle Transporter aktiviert, die Na+-K+-2Cl--Cotransporter. Dem chemischen Gradienten für Na+ folgend, werden auch die anderen 3 Ionen in einem Schritt elektroneutral in die Zelle aufgenommen. Auch hier folgt das Wasser dem Salztransport.
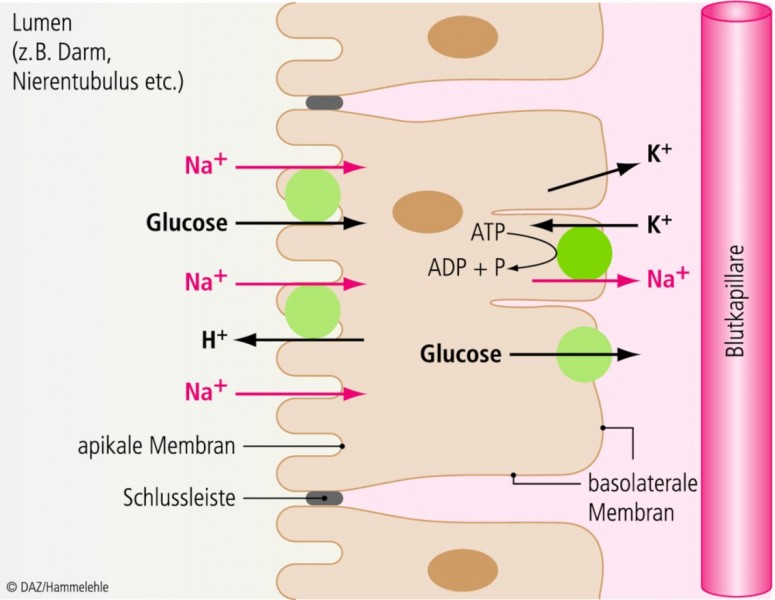
Epitheliale Zellen transportieren geladene Moleküle nicht nur zwischen intra- und extrazellulärem Raum, sondern auch über das Epithel hinweg (transepithelialer Transport). Auch dieser Transport wird durch das Membranpotenzial der Zellen bewerkstelligt. Exokrine Drüsen, Leber oder Niere funktionieren nur mithilfe des Membranpotenzials. Die Grundprinzipien transepithelialen Transports zeigt Abbildung 11. Epitheliale Zellen sind asymmetrisch, sie besitzen eine der Blutseite zugewandte basolaterale Membran, die durch abdichtende Schlussleisten von der apikalen Membran getrennt ist. Über die Schlussleisten sind die Zellen zum Epithel miteinander verknüpft. Die Na+ /K+-ATPase, die immer in der basolateralen Membran zu finden ist, schafft in einem primär aktiven Schritt (d. h. unter Verbrauch von ATP) die für jede Zelle üblichen Gradienten von Na+ und K+. Je nach Transportrichtung werden dann Stoffe wie Glucose, Aminosäuren oder H+ über Cotransporter oder Austauscher (bestimmte Membranproteine) sekundär aktiv im Gefolge von Na+ in die Zelle aufgenommen oder abgegeben. Auch einen epithelialen Na+-Kanal gibt es (ENaC), der aber nicht spannungsabhängig ist. Auf diese Weise können Stoffe aus dem Lumen von Darm oder Nephron resorbiert oder dorthin sezerniert werden.
Abb. 11: Epitheliale Zelle
Transepithelialer Transport, exemplarisch für Ionen und Glucose.
Membranpotenziale in Zellorganellen
Auch innerhalb der Zellen gibt es Kompartimente, die von Membranen umgeben sind, wie z. B. Mitochondrien, das Endoplasmatische Retikulum etc. Auch der Transport über diese intrazellulären Membranen funktioniert nur durch das Potenzial an diesen Membranen. H+-ATPasen schaffen Gradienten über die Membran von sekretorischen Vesikeln. So werden verschiedene Neurotransmitter, z. B. Acetylcholin, in die Vesikel transportiert.
SERCAs (sarko-, endoplasmatisches Reticulum Ca2+-ATPasen) pumpen Ca2+ in intrazelluläre Speicher. Soll es von dort freigesetzt werden, so öffnen Ca2+-Kanäle (IP3- oder Ryanodinrezeptoren). K+-Kanäle in der Membran der Speicher sorgen für Ladungsausgleich, da sonst das Ca2+ durch sein eigenes Nernstpotenzial (= Nullstrompotenzial!) im ER zurückgehalten würde.
Wie bereits oben erwähnt, schaffen die Atmungsketten der Mitochondrien einen H+-Gradienten und damit ein Membranpotenzial über die innere Mitochondrienmembran. Fallen die Protonen über die F1/F0-ATPase zurück in die Matrix, dann läuft diese ATPase quasi "rückwärts" und nutzt den H+-Gradient für die Produktion von ATP.Entkoppelt man die F1/F0-ATPase vom H+-Gradienten, wie das bestimmte H+-Kanäle tun (sogenannte Uncoupling Proteins, UCPs), so entsteht statt ATP bewegungsunabhängig Wärme vor allem im sogenannten Braunen Fettgewebe. Fast alle neugeborenen Säugetiere können sich so vor zu schneller Auskühlung schützen. Zudem haben viele Nagetiere, zum Beispiel Mäuse, so einen eingebauten Ofen, der sie im Winter wärmt (oder zumindest nach dem Winterschlaf schnell aufheizt): Durch den Transport von Ionen mithilfe des Membranpotenzials!
Literatur [1] Bickel-Sandkötter S. Struktur, Funktion und Evolution der ATPasen. Biol Unserer Zeit 1992;3:157 -162.
Autor
Prof. Dr. Peter Krippeit-Drews, Pharmazeutisches Institut, Pharmakologie, Toxikologie und Klinische Pharmazie, Auf der Morgenstelle 8, 72076 Tübingen, peter.krippeit-drews@uni-tuebingen.de



0 Kommentare
Das Kommentieren ist aktuell nicht möglich.