












- DAZ.online
- DAZ / AZ
- DAZ 12/2009
- Wenn Harmlosesgefährlich...
Schwerpunkt Allergie
Wenn Harmlosesgefährlich wird
Kaum werden die Tage wieder länger und die Temperaturen steigen, beginnt die Leidenszeit für einen wesentlichen Teil der Bevölkerung: Die Augen tränen und jucken, die Nase kribbelt und ist verstopft und bei einigen kommt es sogar zu Asthmaanfällen. Und das alles nur, weil Haselnussbüsche und Birkenbäume zu blühen anfangen. Schuld sind Bet v 1, Cor a 1 oder Que a 1 und ähnliche Übeltäter. Hinter diesen Kürzeln verbergen sich die Hauptallergene von Birken-, Haselnuss- und Eichenpollen. Sie gehören, ebenso wie Schimmelpilzsporen, Milbenausscheidungen oder Tierepithelien, zu den Inhalationsallergenen, die also mit der Atemluft an die Schleimhaut des Nasen-/Rachenraums und teilweise bis in die Lungen gelangen. Gemeinsam ist diesen Allergenen, dass es sich um relativ kleine Proteine handelt, die häufig eine enzymatische Aktivität aufweisen. Sie sind im Verhältnis zu anderen Proteinen sehr stabil und können längere Zeit auf trockenen Partikeln wie Milbenkot oder Pollenkörnern ausharren, lassen sich dann an der Schleimhaut aber gut vom Trägerpartikel ablösen und gelangen schließlich in relativ geringer Konzentration in den Mukus. Ein weiteres Charakteristikum ist, dass sie kurze Peptidabschnitte enthalten, die gut über MHC-II präsentiert werden und somit T-Helferzellen stimulieren können. Die Effizienz dieser Präsentation ist allerdings von einer speziellen MHC-II-Ausstattung der betroffenen Personen abhängig, was deutlich macht, dass Allergien – oder allgemein atopische Erkrankungen – eine wichtige genetische Komponente aufweisen.
Die dendritischen Zellen, die sich unter der Haut oder unter den Atemwegsepithelien befinden und dort als erste Abwehreinheit fungieren, haben sich normalerweise als Erste mit eindringenden Parasiten auseinanderzusetzen und sind darauf spezialisiert, Zytokine wie IL-4 auszuschütten, die die Entwicklung der TH 2-Zellen favorisieren. Sie sind es allerdings auch, die des Öfteren auf Bet v 1 und Kollegen treffen und auch diese Proteine aufnehmen, zerkleinern und über MHC-II präsentieren.
Die Allergie
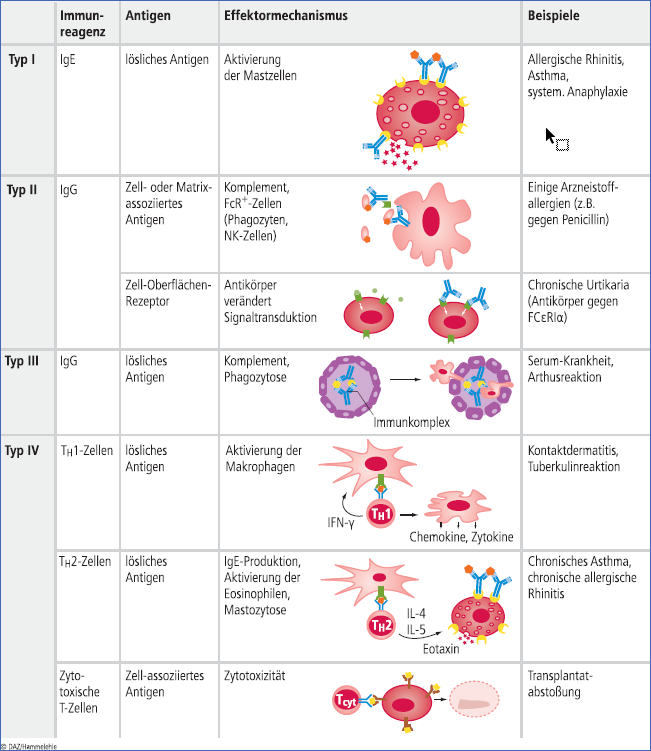
Das, was man landläufig unter Allergie versteht, wird zu den Überempfindlichkeitsreaktionen oder Hypersensitivitätsreaktionen gezählt. Letztlich bedeutet das, dass der adaptive, sehr spezifische Strukturen erkennende Teil unseres Immunsystems – die B-Zellen mit ihren Antikörpern und die T-Zellen – plötzlich auf harmlose Antigene reagiert. Bereits 1963 teilten Robin Coombs und Philip George Houthem Gell die Überempfindlichkeitsreaktionen nach den jeweils zugrunde liegenden Mechanismen in vier verschiedene Typen ein:
Typ I bis III werden von Antikörpern vermittelt, Typ IV von T-Zellen (Abb. 1).
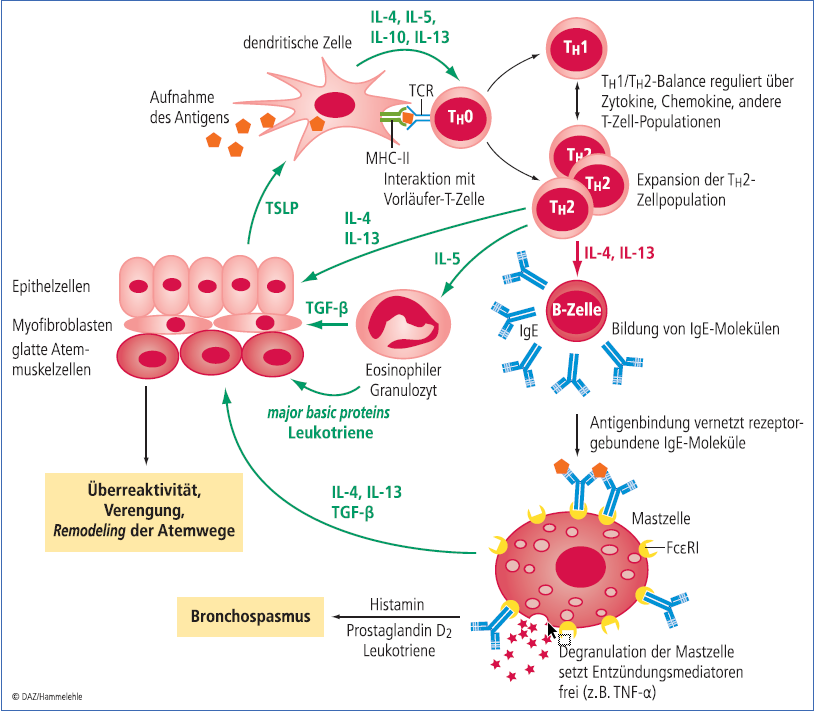
Die Allergie ist eine Typ-I-Reaktion auf die Stimulation von TH 2-Zellen. Diese Untergruppe der T-Helferzellen bildet die Interleukine IL-4 und IL-13 und aktiviert B-Zellen, Antikörper vom IgE-Subtyp gegen das Antigen zu bilden (Abb. 2). IgE-Antikörper sind normalerweise verantwortlich für die Abwehr von Parasiten. Viele dieser Parasiten dringen über Haut und Schleimhaut in den Körper ein, indem sie proteolytische Enzyme ausscheiden, um den festen Zellverband dieser Barriere zu durchdringen. Insofern ist es nachvollziehbar, dass auch die allergenen Enzyme eine IgE-Antwort induzieren.
Nach dem ersten Kontakt mit dem Allergen ist das Immunsystem sensibilisiert und kann bei erneutem Auftreten des Allergens sofort reagieren. Sobald nun aber zwei IgE-Moleküle gleichzeitig ein Allergen und an ihre Rezeptoren (FcεRI) auf einer Mastzelle binden (dies bezeichnet man auch als "bridging"), wird die Mastzelle aktiviert, schüttet ihre Granula aus und setzt dadurch z. B. Histamin, Heparin sowie Enzyme wie Tryptase und Cathepsin G frei (Abb. 2). Im Zuge der Aktivierung bildet die Mastzelle außerdem Zytokine wie IL-3, IL-4, IL-5, IL-13, GM-CSF und TNF-α. Chemokine wie CCL3 und Lipidmediatoren wie die Prostaglandine D2 und E2 , die Leukotriene B4 und C4 sowie der Plättchen-aktivierende Faktor PAF werden neu synthetisiert und ebenfalls sezerniert. Als Folge davon kommt es neben einer umfassenden Entzündungsreaktion zur Kontraktion der glatten Muskulatur, zu einer erhöhten Gefäßdurchlässigkeit und Schleimproduktion – typische Krankheitsbilder z. B. der allergischen Rhinitis oder des Asthmas. Über IL-4 und IL-13 wird der ganze Prozess der TH 2-Zellaktivierung und IgE-Produktion weiter aufrechterhalten. Außerdem mobilisieren IL-3 und IL-5 eosinophile und basophile Granulozyten, die im weiteren Verlauf der allergischen Reaktion über die Ausschüttung z. B. von Enzymen und toxischen Proteinen zu Gewebeschädigungen führen.
Durch die Beteiligung der TH 2-Zellen haben allergische Erkrankungen häufig auch Eigenschaften, die charakteristisch für andere, v. a. Typ-IV-Überempfindlichkeitsreaktionstypen, sind.
Glossar
Granulozyten werden wegen ihrer viellappigen Zellkerne auch als polymorphkernige Leukozyten bezeichnet. Sie enthalten typische Granula im Zytoplasma, die sich je nach Zelltyp mit bestimmten Farbstoffen anfärben lassen, weshalb diese Zellen in neutrophil, eosinophil oder basophil eingeteilt werden.
Eosinophile Granulozyten sind mobile, mäßig phagozytierende Zellen, die vom Blut ins Gewebe migrieren können. Ihre Granula beinhalten toxische Faktoren wie das "major basic protein". Im aktivierten Zustand exponieren die Eosinophilen viele IgE-Rezeptoren auf ihrer Zelloberfläche. Sobald IgE-Moleküle an ihre Rezeptoren binden und über ein Antigen quervernetzt werden, kommt es zur Ausschüttung des Granulainhalts. Eosinophile spielen eine wichtige physiologische Rolle bei der Abwehr von Parasiten.
Mastzellen entwickeln sich im Knochenmark und sind ausschließlich im Gewebe und nicht in der Zirkulation zu finden. Sie tragen hoch affine Rezeptoren für den Fc-Teil von IgE-Antikörpern. Eine Quervernetzung der IgE-Moleküle auf Mastzellen durch Allergenbindung führt zur Degranulation, d. h. zur Freisetzung des Granula-Inhalts mit Proteasen, Histamin etc.
MHC – major histocompatibility complex sind heterodimere Membranproteine, die als Erkennungsmoleküle für T-Lymphozyten dienen. Die MHC-I-Proteine sind auf allen kernhaltigen Zellen vorhanden und werden von zytotoxischen T-Zellen erkannt. Die MHC-II-Proteine kommen vor allem auf antigenpräsentierenden Zellen, einschließlich Makrophagen oder B-Zellen, vor und werden von T-Helferzellen erkannt.
T H 1- und T H 2-Zellen sind Untergruppen der CD4-positiven T-Zellen, die durch spezifische, von ihnen produzierte Zytokine charakterisiert sind. TH 1-Zellen bilden sich in Anwesenheit von IL-12 und Interferon γ (IFN-γ) v. a. nach Stimulation von virus- oder mit intrazellulär persistierenden Bakterien bzw. Protozoen infizierten Makrophagen. Sie produzieren IL-2, IFN-γ, GM-CSF, TNF-α sowie IL-3 und sind an der Aktivierung von zytotoxischen T-Zellen sowie Makrophagen beteiligt. Außerdem stimulieren sie B-Zellen zur Produktion von Antikörpern verschiedener IgG-Subtypen. Sie werden manchmal auch als Entzündungs-T-Zellen bezeichnet. TH 2-Zellen werden durch die Reaktion antigenpräsentierender Zellen auf extrazelluläre Parasiten in Anwesenheit von IL-4 und z.T. auch IL-6 gebildet. Sie produzieren IL-3, IL-4, IL-5, IL-9, IL-10, IL-13 sowie GM-CSF und TGF-β. TH 2-Zellen stimulieren B-Zellen zur Produktion von Antikörpern des IgE-Subtyps.
Strategien gegen Allergien
Verschiedene Strategien werden verfolgt, um der steigenden Zahl von Allergikern zu helfen. Auf eine relativ erfolgreiche Behandlung, der Hyposensibilisierung, also der "Gewöhnung" an das harmlose Antigen, soll hier nicht weiter eingegangen werden. Sie ist Thema des nachfolgenden Beitrags.
Während die Hyposensibilisierung immer Allergen-spezifisch ist, sind die anderen Therapiestrategien unabhängig vom eigentlichen Allergen. Betrachtet man sich den Ablauf der allergischen Reaktion, spielt die Wirkung von Histamin auf H1 -Rezeptoren, z. B. bei der glatten Muskulatur oder auf Endothelzellen, eine entscheidende Rolle. Insofern ist die naheliegende, aber nur palliativ wirkende Therapie die Blockade der H1 -Histaminrezeptoren durch entsprechende Antagonisten. Eine Vielzahl von Wirkstoffen wie substituierte Alkylamine, Cetirizin, Loratadin oder Terfenadin stehen hier zur Verfügung.
Eine Stufe weiter vorne greifen Mastzellstabilisatoren wie die Cromoglicinsäure an. Der postulierte Mechanismus ist eine calciumantagonistische Wirkung, indem Natriumcromoglicat den mit dem IgE-Rezeptor gekoppelten Calciumkanal blockiert. Dadurch wird der Calciumeinstrom in die Mastzelle und somit deren Degranulation verhindert. Es kommt also gar nicht zur Freisetzung von Histamin und anderer Mediatoren. Allerdings tritt der membranstabilisierende Effekt erst nach zwei bis vier Wochen ein.
Bei allen IgE-vermittelten Reaktionen kann auch der Antikörper Omalizumab eingesetzt werden. Omalizumab (Xolair®) ist ein humanisierter rekombinanter, monoklonaler IgG1-kappa-Antikörper, der selektiv an den konstanten Teil des menschlichen Immunglobulin E (IgE) bindet. Dadurch wird verhindert, dass IgE an den FcεRI der Mastzelle bindet, es kann nicht zu einer Quervernetzung der Immunglobuline und somit auch nicht zu einer Aktivierung der Mastzelle kommen. Da Omalizumab alle IgE-Moleküle erkennt und bindet, kann er völlig unabhängig vom eigentlichen, auslösenden Agens angewendet werden. Allerdings ist Xolair® nur als Zusatztherapie zur verbesserten Asthmakontrolle bei Erwachsenen und Jugendlichen (ab 12 Jahren) mit schwerem persistierendem allergischem Asthma zugelassen.
Eine generalisierte Wirkung auf die Zellen des Immunsystems kann mit Glucocorticoiden erreicht werden. Der immunsuppressive Effekt wird über einen intrazellulären Rezeptor realisiert, der zunächst in Form eines Komplexes an ein Hitzeschockprotein gebunden ist. Bindet ein Glucocorticoid an den Rezeptor, dissoziiert das Hitzeschockprotein ab. Der nun in einem aktiven Zustand vorliegende Rezeptor kann einerseits an sogenannte Glucocorticoid-responsive Elemente (GRE) der DNA binden und die Transkription entsprechender Gene initiieren. Er kann aber auch durch Interferenz mit Transkriptionsfaktoren wie AP-1 und NF-κB die Transkription einiger Gene hemmen. Gerade dieser repressive Mechanismus hat besondere Bedeutung für die Suppression der Expression von proinflammatorischen Zytokinen. Insgesamt führt es dazu, dass Glucocorticoide Proliferation, Wachstum und Differenzierung, Adhäsion, Migration und Chemotaxis von Monozyten/Makrophagen, Neutrophilen und T-Zellen hemmen. Zusätzlich werden die Antigenpräsentation durch Monozyten/Makrophagen sowie die Antikörperproduktion von B-Zellen supprimiert und die Apoptose von Monozyten/Makrophagen, T- und B-Zellen gefördert. Wegen dieses allgemein immunsupprimierenden Mechanismus werden Glucocorticoide nicht nur bei schweren Allergien, sondern auch bei anderen Hypersensitivitätsreaktionen angewendet.
Therapien der Zukunft?
Das Wissen um die Zusammenhänge und Interaktionen innerhalb des Immunsystems steigt stetig, wodurch auch immer mehr Zielstrukturen identifiziert werden können, gegen die bei allergischen Reaktionen agiert werden kann.
Wesentliche Schritte in der Aktivierung der Immunzellen bei allergischen Reaktionen werden über die Zytokine IL-4, IL-5 und IL-13 vermittelt. In der Entwicklungsphase befinden sich deshalb Moleküle, die diese Zytokine gezielt "abfangen" wie der lösliche IL-4-Rezeptor Nuvance, die Antikörper gegen IL-4 (Pascolizumab) und gegen IL-5 (Mepolizumab) oder auch ein Fusionsprotein aus einem Teil des IL-13-Rezeptors mit einem konstanten Antikörper-Teil. Die Differenzierung zu TH 2-Zellen lässt sich aber auch dadurch verhindern, dass Zytokine verabreicht werden, die die Immunantwort in Richtung TH 1-Zellen verschieben, wie IL-12, IFN-α oder IFN-γ. Besonderes Augenmerk richtet sich derzeit auf das Thymus-stromale Lymphopoetin (TSLP), das dendritische Zellen in Richtung einer TH 2-vermittelten Entzündungsantwort aktiviert, was wiederum Haut-Keratinozyten zu einer verstärkten TSLP-Expression stimuliert. Könnte man dieses Protein gezielt ausschalten, könnte man ebenfalls eine TH 2-Immunantwort verhindern.
Ziele der späteren Immunantwort sind die eosinophilen Granulozyten, deren Einwanderung an den Ort der allergischen Entzündungsreaktion durch gezielte Blockade beispielsweise des Eotaxin-Rezeptors CCR3 verhindert werden könnte. Ein Antisense-Oligonukleotid, das den CCR3 und auch die gemeinsame β-Kette der Rezeptoren gegen IL-3, IL-5 und GM-CSF blockiert, wird bereits in klinischen Studien gegen allergisches Asthma getestet.
Eine Alternative zu Omalizumab könnte Lumiliximab sein, ein chimärer Antikörper, der gegen den niedrig-affinen IgE-Rezeptor FcεRII (CD23) gerichtet ist und ebenfalls in der Lage ist, die IgE-Serumkonzentration bei Patienten zu reduzieren.
Es bleibt abzuwarten, welche Therapien zukünftig bei allergischen Erkrankungen eingesetzt werden, allerdings werden vermutlich weiterhin bei Heuschnupfen eher die einfacheren, preiswerten Moleküle wie H1 -Antihistaminika als beispielsweise die aufwendigen und teueren Antikörpermoleküle verwendet werden.
Anschrift der Verfasser
Prof. Dr. Angelika Vollmar
LMU München
Department Pharmazie
Pharmazeutische Biologie
Butenandtstr. 5-13
81377 München
Dr. Ilse Zündorf
Prof. Dr. Theo Dingermann
Goethe-Universität Frankfurt
Institut für Pharmazeutische Biologie
Max-von-Laue-Str. 9
60438 Frankfurt
Foto links: foto.fred - Fotolia.com; Foto Logo: emer - Fotolia.com



0 Kommentare
Das Kommentieren ist aktuell nicht möglich.