














- DAZ.online
- DAZ / AZ
- DAZ 19/2015
- CRISPR/Cas9
Genetik
CRISPR/Cas9
Kaum auszusprechen, aber eine Methode mit gewaltigem Potenzial!
Foto: vitstudio - Fotolia.com
Eine merkwürdige Entdeckung
CRISPR ist nicht irgendeine verrückte Erfindung aus einem der unzähligen Genlabors. Es handelt sich vielmehr um ein physiologisches System, das es lange geschafft hat, unentdeckt zu bleiben. Dabei ist es von ungeheurer Wichtigkeit – so wichtig, dass ganz unterschiedliche Organismen dieses Prinzip benutzen. Genauer gesagt handelt es sich um eine Art Immunsystem der Eubakterien und Archae-Bakterien, die sich – ähnlich wie höher entwickelte Lebewesen – auch anstecken können und einen Mechanismus entwickelt haben, um sich gegen diese „Angreifer“ zu wehren. Die „Angreifer“ sind Bakteriophagen – keine Lebewesen also, sondern Informationspakete ähnlich den Viren, die es bekanntlich auf eukaryontische Zellen abgesehen haben.
Im Jahre 1987 publizierten Yoshizumi Ishino und Kollegen von der Osaka University in Japan eine bakterielle Sequenz, die sie iap nannten [1]. Um der Funktion dieser DNA auf die Spur zu kommen, charakterisierten sie auch Genombereiche, die diese iap-Region flankierten. Dabei fanden sie eine Sequenz mit fünf identischen Wiederholungseinheiten, die jeweils 29 Basenpaare enthielten. Diese waren wiederum durch DNA-Blöcke von jeweils 32 Basenpaaren voneinander getrennt. Solche Anordnungen kommen in DNA öfter vor, und man bezeichnet Blöcke, die charakteristische Sequenzmotive voneinander trennen, gemeinhin als Platzhalter bzw. Spacer. Jeder dieser Spacer hatte eine eigenständige Sequenz, ganz anders als die 29-Basen-Sequenzblöcke, die die Spacer voneinander trennten. Die Bedeutung ihrer Entdeckung verstanden Ishino und seine Kollegen damals jedoch noch nicht.
Als um die Jahrhundertwende das Sequenzieren von Genomen unterschiedlicher Mikroorganismen gewissermaßen industrialisiert wurde, entdeckte man in vielen der sequenzierten Organismen – konkret in Eubakterien und Archae-Bakterien – ganz ähnliche Genom-Anordnungen. Da war es Zeit, diesen auffälligen Strukturen einen Namen zu geben, und so „taufte“ der Niederländer Ruud Jansen von der Universität in Utrecht im Jahre 2002 diese Genom-Konstellation als clustered regularly interspaced short palindromic repeats (CRISPR). Zusätzlich machte die holländische Gruppe eine weitere wichtige Entdeckung: Den Wissenschaftlern fiel nämlich auf, dass sich in direkter Nachbarschaft zur CRISPR-Region Gene befanden, die sie als CRISPR-associated genes (Cas) bezeichneten.
Eine interessante Abwehrstrategie
Es dauerte noch ein paar Jahre, bis bei Sequenzvergleichen auffiel, dass die Spacer-Sequenzen verblüffende Übereinstimmungen mit Phagen-Sequenzen zeigten. Und so wurde recht schnell die Hypothese geboren, dass das CRISPR/Cas-System eine weitere, bisher unbekannte Variante eines bakteriellen Abwehrsystems – quasi ein primitives Immunsystem – repräsentieren könnte.
Könnte es sein, so die Hypothese, dass die Cas-Enzyme bei einer Infektion eines Bakteriums mit einem Bakteriophagen einen Teil der Phagen-DNA ausschneiden und diese dann in ihre CRISPR-Region inserieren würden? Damit wäre gewissermaßen ein Infektionsgedächtnis etabliert, quasi ein Analogon zu unserem spezifischen Immunsystem, das bei einer erneuten Infektion von Nutzen sein könnte. Dass sich Bakterien gegen Fremd-DNA wehren können, ist längst bekannt. Eines der prominentesten Beispiele sind die Restriktionsenzyme, die eine Phagen-DNA dadurch erkennen, dass sie im Gegensatz zur bakteriellen DNA an den Erkennungssequenzen für die Restriktionsendonuklease nicht modifiziert ist und folglich geschnitten wird. Im Falle von CRISPR/Cas ist die Situation ähnlich, aber dann doch auch deutlich anders.
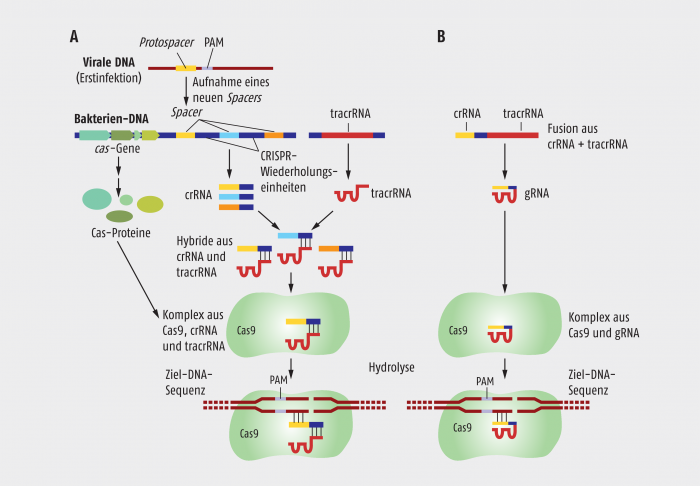
In Bakterien, die ein CRISPR/Cas-System besitzen, fungiert die Cas-Nuklease als eine Art Präzisionsschere. Diese bildet einen Komplex mit einem Molekül RNA (crRNA), das von einem
Spacer-CRISPR-DNA-Segment abgelesen wurde (Abb. 1A). Findet diese crRNA eine komplementäre Sequenz auf der DNA eines eingedrungenen Phagen, so bindet sie spezifisch und fest an die Phagen-DNA, woraufhin die Cas-Nuklease die DNA hinter dem DNA/RNA-Heteroduplex zerschneidet.
Grafik: I. Zündorf
Abb. 1: Natürlich vorhandene und synthetisch hergestellte CRISPR-Cas-Systeme. A. Beim natürlich vorkommenden CRISPR-System werden fremde DNA-Sequenzen in sogenannten CRISPR-Arrays in das Genom der Wirtszelle eingebaut. Von diesen Bereichen werden CRISPR-RNA-Stücke (crRNAs) mit Sequenzabschnitten transkribiert, die komplementär zur DNA des Eindringlings sind. Erst nach Hybridisierung mit einer anderen, vom CRISPR-System codierten RNA, der transaktivierenden CRISPR-RNA (tracrRNA), kann ein Komplex mit der Cas9-Nuklease ausgebildet werden, der dann wiederum die Fremd-DNA, die an die Spacer-Sequenz gebunden hatte, schneidet. Im Unterschied zur genomischen Spacer-Sequenz der Wirtszelle befindet sich auf der Fremd-DNA die sogenannten PAM-Sequenz (protospacer adjacent motif) in unmittelbarer Nähe der Erkennungssequenz. B. Für die Verwendung im Labor wurde eine crRNA-Sequenz mit einem Teil der tracrRNA fusioniert. Nach der Transkription dieser Informationseinheit erhält man eine gRNA, die einerseits einen Komplex mit Cas9 bilden kann, andererseits einen Sequenzbereich zur Erkennung einer Ziel-DNA trägt.
Von CRISPR-RNA gesteuerte Endonuklease Cas9 – Ein komplexer Mechanismus …
Um den spezifischen Zerstörungsmechanismus gegen eingedrungene Phagen in Gang zu bringen, werden zunächst Transkripte von CRISPR-Blöcken zu CRISPR-RNAs prozessiert (crRNAs). Jede dieser RNAs besteht aus einer variablen Sequenz, der „Spacer“-Sequenz, die von einem Phagen stammt, der einmal die Zelle infiziert hatte, und aus einem Teil der identischen CRISPR-Wiederholungseinheiten. Die crRNAs hybridisieren mit einer zweiten RNA, der sogenannten transaktivierenden CRISPR-RNA (tracrRNA). Diese beiden RNAs bilden dann schließlich einen Komplex mit der Cas-Nuklease.
Infiziert ein Phage eine mit einem CRISPR/Cas-System geschützte Zelle, so dirigiert die Spacer-Sequenz in der crRNA die Cas-Nuklease zu der komplementären Phagen-DNA. Befindet sich neben dem 5’-Ende der Protospacer-Sequenz eine sogenannte PAM-Sequenz (protospacer adjacent motif) auf der Phagen-DNA, hydrolysiert die Cas-Endonuklease an dieser Stelle die DNA des Eindringlings. Die Spacer-Sequenzen, die ja auch im CRISPR-Locus auf dem Bakterien-Genom vorhanden ist, wird nicht zerstört, da sie sich nicht neben einer PAM-Sequenz befindet.
… und seine Vereinfachung für die Praxis
Das am weitesten eingesetzte CRISPR/Cas-System ist das CRISPR/Cas9-System aus Streptomyces pyogenes. Dieses System wurde für ein gezieltes Genom-Editing in vielfältiger Weise angepasst. In seiner einfachsten und am weitesten verbreiteten Form werden zwei Komponenten verwendet, die in den Organismus eingebracht werden müssen, dessen DNA punktgenau verändert werden soll. Dies ist einmal das Gen für die Cas9-Endonuklease und zum anderen eine sogenannte Führungs-RNA (guide RNA/gRNA). Diese besteht aus einer Fusion einer speziell ausgewählten crRNA und einer konstanten tracrRNA (Abb. 1B). Die Cas9-Nuclease spaltet die Ziel-DNA dann, wenn die Protospacer-DNA an ihrem 5’-Ende neben einer PAM-Sequenz liegt. Diese besitzt in der üblichen Form die Sequenz 5’-NGG. Man kann sich leicht vorstellen, wie flexibel dieses System einsetzbar ist. In der Ziel-DNA muss man nur eine 5’NGG-PAM-Sequenz identifizieren und von da aus eine komplementäre Spacer-RNA synthetisieren, der dann die konstanten Bereiche der gRNA folgen.
Zur spezifischen Spaltung kommen Mechanismen der DNA-Reparatur
Um das CRISPR/Cas-System als Genom-Editierungssystem voll verstehen zu können, gilt es noch eine wichtige Komponente zu betrachten: Die Korrektur von Doppelstrangbrüchen. In Abwesenheit exogener homologer DNA- oder RNA-Reparatur-Matrizen können durch Doppelstrangbrüche, wie sie von der Cas-Endonuklease generiert werden, Insertionen oder Deletionen (sogenannte
Indels) über eine fehlerhafte End-zu-End-Verknüpfung (error-prone nonhomologous end-joining/NHEJ) hervorgerufen werden. Durch homologe Rekombination (HR) mithilfe einer Reparatur-Matrize kann die zu reparierende DNA gezielt verändert werden (Abb. 2).
Grafik: I. Zündorf
Abb. 2: Genom-Modifikation über DNA-Reparaturmechanismen. Ein DNA-Doppelstrangbruch (DSB) wird üblicherweise entweder über eine nicht-homologe Verknüpfung der entstandenen Enden (nonhomologous end-joining, NHEJ) oder anhand einer homologen DNA-Sequenz (homology-directed repair, HDR) repariert. Die NHEJ ist sehr fehleranfällig und führt durch Insertionen bzw. Deletionen von Nukleotiden (Indel-Mutationen) eventuell zu Leserasterverschiebungen oder falschen Stopp-Codons. Bei der HDR wird beispielsweise das Schwesterchromatid verwendet, um den Doppelstrangbruch zu reparieren. Bietet man hier eine exogen eingefügte DNA zur Reparatur an, lässt sich gezielt eine DNA-Modifikation durchführen.
Die bemerkenswerte Vielfalt der Anwendungsmöglichkeiten des CRISPR/Cas-Systems
Nachdem im Jahre 2009 erstmals die Flexibilität dieses ausgeklügelten bakteriellen Immunsystems gezeigt worden war, folgte eine große Zahl von Publikationen, in denen die Anwendung in allen möglichen Organismen demonstriert wurde. Dies betraf nicht nur die Genome von Mikroorganismen und Zellkulturzellen, sondern auch die Genome ganzer Organismen, darunter u. a. Hefe, Tabak, Reis, Weizen, Maus, Ratten, Kaninchen, Frosch, Drosophila, Zebrafisch und auch das menschliche Genom ließen sich spezifisch editieren. Die Möglichkeiten der genetischen Veränderungen sind zudem extrem vielfältig. So kann man bestimmte Sequenzen an bestimmten Stellen eliminieren. Man kann aber auch Sequenzen einfügen oder DNA-Regionen beispielsweise mit leuchtenden Proteinen für die Mikroskopie markieren. Schließlich lassen sich auch Genomregionen aktivieren, die in bestimmten Zellen inaktiv vorliegen. Dazu wurden u. a. spezielle Varianten der Cas9-Endonuklease entwickelt, die bei ganz bestimmten Problemstellungen zu sehr differenzierten Lösungen führen können. Diese unglaublich schnelle Verbreitung des CRISPR/Cas-Systems wurde auch dadurch möglich, dass viele Komponenten der Cas9-Technologie durch open-source-Distributoren wie Addgene zu Verfügung gestellt wurden und dass sich Anwender in Foren wie www.genome-engineering.org oder www.egenome.org rege austauschten.
From Bench to Bedside
Normalerweise wäre es vermessen, bei einer so jungen Technologie bereits jetzt schon nach der Translation in die Klinik zu fragen, obwohl sich nicht nur die Technologievariationen, sondern auch die Fantasien in Richtung klinischer Anwendung überschlagen. Dennoch sollten wir uns nicht wundern, wenn bald schon konkrete Berichte auftauchen, die erste Anwendungen beim Menschen schildern. Die spektakulärste – und gleichzeitig auch erschreckendste – Meldung in dieser Hinsicht konnten wir kürzlich von chinesischen Forschern lesen, die erstmals eine Gentherapie unter zu Hilfenahme des CRISPR/Cas-Systems bei menschlichen Embryonen wagten. Dies ist erschreckend deshalb, da es hier – sollte die Meldung sich als valide erweisen – zu einem ethischen Dammbruch gekommen wäre. Denn eine Gentherapie an Keimbahnzellen galt bisher als allgemein nicht akzeptabel. Vergleichsweise harmlos, wenn auch nicht weniger spektakulär, ist da der Vorschlag des Harvard-Kardiologen Kiran Musunuru, der mithilfe der CRISPR/Cas9-Technologie das Gen für das Leberenzym PCSK9 zerstören will, um so das Risiko für Herz-Kreislauf-Erkrankungen um deutlich über 60% zu senken [2]. Dies leitet Musunuru aus Mausstudien ab, die die Gruppe im letzten Jahr in der Zeitschrift Circulation Research publizierten [3].
Grafik: I. Zündorf
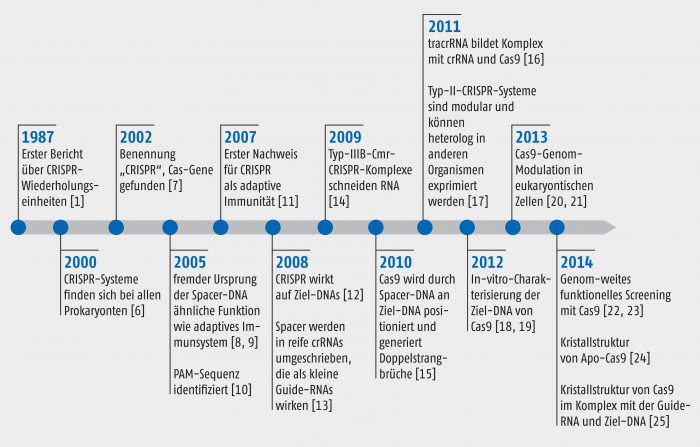
Abb. 3: Zeittafel wichtiger Entdeckungen und Optimierungen des CRISPR-Systems
40 Jahre nach Asilomar: das Napa-Meeting
1975, zu Beginn der Gentechnik-Ära, trafen sich Wissenschaftler, um Richtlinien zur Produktion und Handhabung gentechnisch veränderter Organismen festzulegen. Diese Richtlinien wurden von zahlreichen Ländern übernommen und waren später die Basis für das Deutsche Gentechnikgesetz. Im Januar 2015 fand ein ähnliches Treffen im kalifornischen Napa statt, um eine öffentliche Diskussion über die Möglichkeiten der Genom-Modifikationen mit Methoden wie z. B. das CRISPR/Cas9-System anzustoßen. Gerade angesichts der Tatsache, dass die Erforschung des CRISPR/Cas9-Systems so rasant vorangeschritten ist (Abb. 3) und dass eventuell chinesische Wissenschaftler bereits menschliche Embryonen genetisch manipuliert haben, ist eine weltweite und kritische Diskussion über Risiken und Verheißungen dieser Technologie dringend nötig. Die Wissenschaftler der Napa-Konferenz, die teilweise auch schon vor 40 Jahren in Asilomar mit dabei waren, warnen vor einer zu schnellen Anwendung des Systems am Menschen, fordern aber gleichzeitig die Unterstützung der transparenten Erforschung der Genom-Modifikationen, um mögliche Risiken besser abschätzen zu können [4, 5]. |
CRISPR/Cas9 – was braucht man dafür?
Mittlerweile sind bei verschiedenen Anbietern für molekularbiologische Werkzeuge auch entsprechende Klonierungsvektoren für das CRISPR-System verfügbar. Was ein derartiges Plasmid typischerweise enthält, ist in der Abbildung zu sehen.
Um die DNA in Bakterien vervielfältigen zu können, muss das Plasmid einen Replikationsursprung (origin of replication, Ori) tragen, an dem die Verdopplung der DNA startet (hier: pUC-Ori), sowie ein Resistenzgen für die Selektion der Bakterienklone, die die DNA aufgenommen haben (hier: Resistenz gegenüber Kanamycin, KanR). Da dieses Plasmid in Säugerzellen extrachromosomal repliziert werden soll, ist zudem der Ori des Simian-Virus 40 (SV40-Ori) nötig. Ein essenzieller Bestandteil des CRISPR-Systems ist die Cas9-Nuklease, die unter der Kontrolle eines Promotors exprimiert wird. Je nachdem, wo und wie das System angewendet werden soll, bieten sich hier verschiedene, jeweils sehr spezifische Promotorsequenzen an. Damit die Nuklease auch wirklich in den Zellkern gelangt, sind zwei Kernlokalisierungssequenzen (nucleus localisation signal, NLS) mit dem Gen fusioniert. Als Markierung der Nuklease bzw. der transfizierten Zelle kann zudem beispielsweise ein kurzes Peptid (myc-tag) oder aber ein fluoreszierendes Protein dienen. Außerdem hilft ein weiteres Sequenzelement, das Woodchuck virus post-transcriptional regulatory element (WPRE), die Expression des Cas9-Gens noch zu steigern und die mRNA zu stabilisieren.
Grafik: Zündorf
Abb. 4: Vektorkarten für ein CRISPR-System
Ein weiterer, sehr wichtiger Bereich ist die gRNA-scaffold-Sequenz. Hier befindet sich bereits die Information für die tracrRNA und es kann eine beliebige, kurze DNA eingesetzt werden, die zur Ziel-DNA im Genom oberhalb einer NGG3’-PAM-Sequenz komplementär ist. Dorthin wird nach der Transfektion der Komplex aus RNA und Nuklease gelenkt, um einen Doppelstrangbruch zu generieren.
Wird gleichzeitig ein Donor-Plasmid mit transfiziert, kann über die beiden Rekombinationsarme mit Sequenzen, die komplementär zu den Ziel-Gensequenzen neben der Cas9-Schnittstelle sind, eine Reparatur der Bruchstelle durchgeführt werden. Dabei wird das Donor-Plasmid in das Genom integriert, wobei mittels einer erworbenen Antibiotika-Resistenz (hier: Puromycin, PuroR) auf das Integrationsereignis selektioniert werden kann. Befinden sich außerdem noch loxP-Sequenzen auf dem Donor-Plasmid, lässt sich der dazwischen liegende DNA-Bereich mit dem Antibiotika-Resistenzgen wieder entfernen.
Literatur
[1] Ishino et al. (1987): Nucleotide Sequence of the iap Gene, Responsible for Alkaline Phosphatase Isozyme Conversion in Escherichia coli, and Identification of the Gene Product. Journal of Bacteriology, 1169,5429-5433
[2] Colen BD. A shot against heart attacks. http://news.harvard.edu/gazette/story/2014/06/a-shot-against-heart-attacks/
[3] Ding et al. (2014): Permanent Alteration of PCSK9 With In Vivo CRISPR-Cas9 Genome Editing. Circulation Research 115;488-492
[4] Vogel (2015): Embryo engineering alarm. Science 347,1301
[5] Baltimore et al. (2015): A prudent path forward for genomic engineering and germline gene modification. Science 348,36-38
[6] Mojica et al. (2000): Biological significance of a family of regularly spaced repeats in the genomes of Archaea, Bacteria and mitochondria. Mol. Microbiol. 36,244-246
[7] Jansen et al. (2002): Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 43,1565-1575
[8] Mojica et al. (2005): Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements. J. Mol. Evol. 60,174-182
[9] Pourcel et al. (2005): CRISPR elements in Yersinia pestis acquire new repeats by preferential uptake of bacteriophage DNA, and provide additional tools for evolutionary studies. Microbiology 151,653-663
[10] Bolotin et al. (2005): Clustered regularly interspaced short palindrome repeats (CRISPRs) have spacers of extrachromosomal origin. Microbiology 151,2551-2561
[11] Barrangou et al. (2007): CRISPR provides acquired resistance against viruses in prokaryotes. Science 315,1709-1712
[12] Marraffini et al. (2008): CRISPR interference limits horizontal gene transfer in staphylococci by targeting DNA. Science 322,1843-1845
[13] Brouns et al. (2008): Small CRISPR RNAs guide antiviral defense in prokaryotes. Science 321,960-964
[14] Hale et al. (2009): RNA-guided RNA cleavage by a CRISPR RNA-Cas protein complex. Cell 139,945-956
[15] Garneau et al. (2010): The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature 468,67-71
[16] Deltcheva et al. (2011): CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature 471,602-607
[17] Sapranauskas et al. (2011): The Streptococcus thermophilus CRISPR/Cas system provides immunity in Escherichia coli. Nucleic Acids Res. 39,9275-9282
[18] Jinek et al. (2012): A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337,816-821
[19] Gasiunas et al. (2012): Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc. Natl. Acad. Sci. USA 109, E2579-E2586
[20] Cong et al. (2013): Multiplex genome engineering using CRISPR/Cas systems. Science 339,819-823
[21] Mali et al. (2013): RNA-guided human genome engineering via Cas9. Science 339,823-826
[22] Wang et al. (2014): Genetic screens in human cells using the CRISPR-Cas9 system. Science 343,80-84
[23] Shalem et al. (2014): Genome-scale CRISPR-Cas9 knockout screening in human cells. Science 343,84-87
[24] Jinek et al. (2014): Structures of Cas9 endonucleases reveal RNA-mediated conformational activation. Science 343,1247997
[25] Nishimasu et al. (2014): Crystal structure of Cas9 in complex with guide RNA and target DNA. Cell 156,935-949
Autoren
Prof. Dr. Theo Dingermann ist Seniorprofessor am Institut für Pharmazeutische Biologie an der Goethe-Universität Frankfurt.
Dr. Ilse Zündorf ist dort als akademische Oberrätin tätig.
Institut für Pharmazeutische Biologie
Biozentrum
Max-von-Laue-Straße 9
60438 Frankfurt/Main



0 Kommentare
Das Kommentieren ist aktuell nicht möglich.