















- DAZ.online
- DAZ / AZ
- DAZ 50/2001
- Bioverfügbarkeit von ...
Spurenelemente
Bioverfügbarkeit von Zinkpräparaten
Fleischkonsum und Zinkstatus
Das Deutsche Institut für Ernährungsmedizin und Diätetik (DIET e.V.) warnte unlängst vor einer Mangelversorgung mit essenziellen Nährstoffen, ausgelöst durch eine plötzliche Änderung der Ernährungsgewohnheiten [1]. Aufgrund der aktuellen Krisen in der Landwirtschaft (BSE, Maul-undKlauen-Seuche, Massentierhaltung, Antibiotika und andere Arzneimittel als Masthilfen) ist in der Bevölkerung ein rückläufiger Fleischkonsum zu beobachten, der sich auch auf die Versorgung mit essenziellen Nährstoffen auswirken kann. Von diesen Entwicklungen vor allem betroffen ist das Spurenelement Zink.
Der Zinkstatus bei Menschen wird in erster Linie von drei Faktoren beeinflusst [2]:
- Ausmaß der Zinkzufuhr,
- gesteigerte Zinkverluste oder höherer Zinkbedarf bei bestimmten Erkrankungen wie Diabetes, chronischen Entzündungen und dermatologischen Erkrankungen,
- gesteigerte Zufuhr von Inhibitoren der Zinkaufnahme mit der Nahrung.
Fleisch ist unsere Hauptquelle für die Zinkzufuhr. Der plötzliche Verzicht auf Fleisch geht zumeist mit einer Erhöhung der Zufuhr vegetabiler Nahrung einher und somit einer erhöhten Zufuhr von Phytaten, was die Zinkversorgung zusätzlich negativ beeinflusst.
Besonders für Risikogruppen, zu denen neben Kranken auch schwangere und stillende Frauen, Kinder in Wachstumsphasen, Senioren, Rekonvaleszente und Sportler zählen, empfiehlt sich die Substitution von Zink mit einem als Arzneimittel zugelassenen und in seiner pharmazeutischen Qualität gesicherten Zinkpräparat. Wenig bekannt ist bislang, dass sich Zinkpräparate je nach eingesetzter Zinkform teilweise erheblich in ihrer Bioverfügbarkeit unterscheiden.
Problematische Bioverfügbarkeit
Die Bioverfügbarkeit von Zink variiert beträchtlich. Insbesondere pflanzliche Kost ist zumeist eine schlechte Zinkquelle. Zink bildet mit den in Pflanzen enthaltenen Ballaststoffen, vor allem der Phytinsäure, schwerlösliche Komplexe, die einer Resorption entzogen sind. Verantwortlich für diesen Effekt sind insbesondere Phytate vom Typ des Inositolpenta- bzw. -hexaphosphates. Die Zufuhr von Zink kann demzufolge auch dann ungenügend sein, wenn Analysen der pflanzlichen Kost ausreichende Zinkgehalte andeuten [2]. Auch Oxalat oder Tannine [4] sowie Schwermetalle wie Cadmium [2] können die Zinkaufnahme behindern. Die Beeinflussung durch Cadmium dürfte in der Praxis allerdings keine relevante Rolle spielen.
Problematische Resorption
Die Zufuhr der empfohlenen Tagesdosis von 15 mg
Zink kann dann unzureichend sein, wenn aufgrund
der schlechten Bioverfügbarkeit des Mineralstoffs
aus der jeweiligen Zinkquelle nur Bruchteile dieser
Zinkdosis zur Resorption kommen [3].
In der medizinischen Literatur wird eine gegenseitige Resorptionsbeeinflussung verschiedener Mineralstoffe diskutiert. Eine postulierte Interaktion mit Calcium erschien unter verschiedenen Versuchsbedingungen eher zweifelhaft [2]. Vergleichbares gilt für die Interaktion zwischen Kupfer und Zink [2]. Trotzdem wird bei gleichzeitiger Einnahme von Zink und Kupfer vor sekundärem Kupfermangel gewarnt [5]. Die Relevanz dieser Wechselwirkung ist eher zweifelhaft [3]. So beobachteten Sullivan et al. [6] im Rahmen ihrer Bioverfügbarkeitsstudie mit Zinksulfat keinerlei Beeinflussung des Kupfer-, Calcium- und Magnesiumspiegels trotz Verabreichung der relativ hohen Dosis von 50 mg Zink in einer Einmaldosis. Dennoch empfiehlt es sich, die Einnahme von Kupfer-Supplementen zeitlich von der von Zinksupplementen zu trennen.
Hemmung der Zinkaufnahme durch Eisen?
Auch die bei gleichzeitiger Gabe von Eisen und Zink beobachtete Hemmung der Zinkresorption tritt möglicherweise nur unter den artifiziellen Bedingungen des Probandenversuchs auf. Solomons und Jacob [7] stellten fest, dass hochdosiertes anorganisches Eisen die Zinkaufnahme hemmte, gemessen an der Veränderung des Plasma-Zinkspiegels innerhalb von vier Stunden nach oraler Einnahme von Zink. Die Untersucher verabfolgten an erwachsene, nüchterne Probanden jeweils 25 mg Zink in Form von Zinksulfat und Eisen in einer Dosis von 25, 50 oder 75 mg. Bereits bei 25 mg Eisen war die Zinkaufnahme signifikant niedriger als bei den Kontrollpersonen.
Unter physiologischeren Bedingungen scheint dieser Effekt jedoch weniger stark ausgeprägt zu sein [8] und bei Zugabe von Histidin zur Testdosis sogar dann aufgehoben, wenn das Dosisverhältnis zwischen Eisen und Zink mit 25 : 1 in unphysiologischen Bereichen lag. Dieses Ergebnis wurde mittlerweile in verschiedenen tierexperimentellen und klinischen Untersuchungen mit Radioisotopen bestätigt (z. B. [9, 10]).
Eisen und Zink können in pharmazeutischen Zubereitungen bedenkenlos gemeinsam eingesetzt werden [2]. Auch die Langzeiteinnahme von Eisen- und/oder Zinksupplementen hat keinen negativen Einfluss auf den Versorgungsstatus beider Mineralstoffe [2, 8].
Bessere Zinkresorption durch Kopplung an Aminosäuren
Gute Zinkquellen sind zumeist tierische Produkte, zum Beispiel Rindfleisch oder Milchprodukte. Die Proteine in der Nahrung beeinflussen die Zinkaufnahme, wobei verschiedene Proteine durchaus unterschiedliche Effekte zur Folge haben können. So ist zum Beispiel für Casein eine geringgradige Hemmung der Zinkaufnahme bekannt.
Dagegen zeigen bestimmte Aminosäuren, insbesondere Methionin, Cystein oder Histidin, einen deutlich fördernden Einfluss auf die Zinkaufnahme [2]. Der positive Einfluss von Nahrungsproteinen tierischen Ursprungs wird denn auch weniger auf den Einfluss des Eiweißmoleküls selbst als vielmehr auf die daraus freigesetzten freien Aminosäuren zurückgeführt. Diese Aminosäuren können Zink auch in Anwesenheit von Phytaten in gelöster, resorbierbarer Form halten - ein Effekt, der auch für Oligopeptide aus Casein bereits nachgewiesen wurde.
Muttermilch als Zinkquelle
Hinsichtlich der Bioverfügbarkeit von Zink bestehen zwischen verschiedenen Nahrungsmitteln erhebliche Unterschiede. Da Kinder in Wachstumsphasen sensibel auf die Zinkversorgung reagieren, ist die Beachtung des Zinkstatus in der Kinderernährung besonders wichtig. Für Säuglinge ist die Muttermilch die beste Zinkquelle - vorausgesetzt, die Mutter weist einen ausreichenden Zinkstatus auf. So wurde anhand radioaktiv markierter Zinkzusätze in tierexperimentellen Studien an Ratten festgestellt, dass die Zink-Bioverfügbarkeit aus menschlicher Muttermilch bei 28% liegt, aus Kuhmilch bei 15% und aus Soja-Formula-Nahrung bei nur 10% [11].
Evans et al. machten eine Komplexbildung mit Picolinsäure für die gute Bioverfügbarkeit von Zink aus Muttermilch verantwortlich [12]. Während Picolinsäure, die mit Zink einen Chelatkomplex bilden kann, in der Muttermilch in einer Konzentration von 308 Mikromol/l vorkommt, beträgt die Konzentration in Kuhmilch lediglich 20 Mikromol/l oder weniger. Picolinsäure verbessert die Bioverfügbarkeit von Zink: Barrie et al. [13] fanden im Rahmen eines vierwöchigen Probandenversuchs eine Erhöhung des Zinkspiegels in den Haaren, im Urin und den Erythrozyten.
Allerdings reicht einigen Autoren zufolge die Konzentration von Picolinsäure in der Muttermilch nicht aus, um die Verfügbarkeit von Zink aus dieser Quelle befriedigend zu erklären. So weist Zinkpicolinat gegenüber dem Komplex Zink-Histidin eine deutlich unterlegene Verfügbarkeit auf [14-16]. Histidin als essenzielle Aminosäure ist gleichfalls Bestandteil der Muttermilch und könnte einen zusätzlichen Beitrag für die gute Bioverfügbarkeit von Zink in Muttermilch leisten [11].
Resorptionsverbesserer für Zink
Verschiedene niedermolekulare Substanzen wurden auf ihre resorptionsfördernde Wirkung für Zink untersucht:
- Ascorbinsäure, deren positiver Einfluss auf die Eisenaufnahme gut bekannt ist, bewirkte in Dosen zwischen 0,5 und 2 Gramm nach Gabe von 110 mg Zinksulfat-Heptahydrat, entsprechend 25 mg Zink, keine Veränderung der Zinkaufnahme [17].
- Auch für Citronensäure waren die Ergebnisse unbefriedigend. Der Zusatz von Citrat zu Lebensmitteln kann zwar unter Umständen die Resorption von Zink verbessern [18]. Für eine kontrollierbare Supplementation mit Zinkpräparaten kommt Zinkcitrat jedoch nicht infrage. Barrie et al. [13] konnten zudem bei Gabe von Zinkcitrat keine Veränderung des Serum-Zinkspiegels nachweisen.
- EDTA (Ethylendiamintetraacetat), eine Chelat-bildende Substanz, die zur Komplexierung von Metallionen eingesetzt wird, wurde bereits frühzeitig als potenzielle Zinkfähre in Tierversuchen an Hühnern getestet [19, 20]. Allerdings stellte sich in der Folge heraus, dass komplexiertes Zink-EDTA zwar gut aus dem Darmlumen in die Enterozyten aufgenommen wird, von hier aus aber nicht die basolaterale Membran durchdringen kann [21]. Um für den Organismus verwertbar zu sein, müssen die Zinkionen daher zunächst umkomplexiert werden.
- Aminosäuren sind gute Zink-Chelatoren, insbesondere Histidin [2, 5, 22, 23]. Die Eignung von Zink-Histidin als Zinklieferant wurde in klinischen Studien bewiesen (s. u.). Dabei kommt es auf das Mengenverhältnis zwischen Zink und Histidin an: Extrem hohe Mengen an Histidin (60 Gramm pro Tag), die weder in der Nahrung noch in Zinksupplementen vorkommen, können sogar zu einer verstärkten Ausscheidung von Zink über den Urin führen [24]. Zink-Histidin wird jedoch bei hoher Dosierung renal ausgeschieden [25], was eine - beabsichtigte oder versehentliche - Überdosierung ausschließt.
Zinkformulierungen in Handelspräparaten
- Zinkoxid
- Zinksulfat
- Zinkgluconat
- Zinkorotat
- Zinkaspartat
- Zink-Histidin
Zinkformulierungen
Zinkpräparate des Handels enthalten die Zinkionen zumeist in Form anorganischer Salze oder organischer Salze oder in Komplexen mit Aminosäuren (s. Kasten). Für diesen Beitrag wurden mehr als 500 einschlägige Berichte aus der medizinischen Literatur gesichtet. Aussagekräftige klinische Untersuchungen liegen dabei offensichtlich nur für wenige Zinkformen vor. Insbesondere für den Komplex aus Zink und Histidin wurden im Vergleich zu Zinksulfat relevante Vorteile hinsichtlich der Bioverfügbarkeit klinisch nachgewiesen. Zinksulfat diente in den meisten Studien als Referenzpräparat.
Zinkcarbonat
Zinkcarbonat ist kein üblicher Bestandteil von Zinksupplementen [31]. Es ist schlecht löslich und keine gute Quelle für die Resorption von Zink [5].
Zinksulfat
Zinksulfat kann für die vergleichende Betrachtung der Bioverfügbarkeit als Referenzpräparat gelten. In einer Reihe von Studien wurde mit diesem Arzneistoff die Abhängigkeit der Zinkresorption von verschiedenen Faktoren untersucht. Ein Faktor, der die Aufnahme von Zinksulfat eindeutig negativ beeinflusst, ist die Einnahme unmittelbar zu den Mahlzeiten. So fanden Keyzer et al. keine erhöhten Serum-Zinkwerte bei Zufuhr von 50 mg Zink in Form von Zinksulfat, wenn das Zinkpräparat zum Essen gegeben wurde, aber eine Erhöhung um den Faktor 2 bei Nüchterneinnahme [32].
Sullivan et al. (1979) fanden bei Nüchterneinnahme von 220 mg Zinksulfat, entsprechend 50 mg Zink, nicht nur eine Erhöhung der Serum-Zinkspiegel, sondern auch eine 140%ige Steigerung der renalen Zinkausscheidung [6]. Pohit et al. (1981) und Solomons (1982) fanden gleichfalls einen signifikanten Anstieg der Serum-Zinkwerte nach Nüchterneinnahme von Zinksulfat [3, 33].
Zinkoxid
Zinkoxid ist in der Dermatologie als Bestandteil externer Zubereitungen bekannt, zum Beispiel in Wund- und Heilsalben. Es ist aber auch als Wirkstoff für die orale Supplementation gebräuchlich, insbesondere in Nahrungsergänzungsmitteln ohne amtliche Zulassung.
Wedekind et al. stellten im Tierversuch an Hühnern die relativen Unterschiede der Bioverfügbarkeit verschiedener Zinkformen im Vergleich zu Zinksulfat (100%) fest [26]. Demnach hatte Zinkoxid eine relative Bioverfügbarkeit von nur von 61%. Umgerechnet auf die Einlagerung von Zink in die Knochen der Versuchstiere, also die Verwertung des zugeführten Zinks, schnitt Zinkoxid noch schlechter ab: mit 44% des Wertes von Zinksulfat [27].
Henderson et al. (1995) verglichen an zehn gesunden Probanden die Plasma-Zinkkonzentrationen nach einmaliger Gabe von Zinkoxid bzw. Zinkacetat. Diese Autoren bezeichnen Zinkoxid vor allem bei Patienten mit Störungen der Magensäureproduktion als eine ungeeignete Zinkverbindung [30].
Zinkgluconat
Für eine abschließende Bewertung von Zinkgluconat als Zinkquelle sind noch einige Fragen offen. So fanden Barrie et al. [13] in einer Probandenstudie an 15 gesunden Freiwilligen bei jeweils 4-wöchiger, doppelblinder Verabreichung von Zinkgluconat, Zinkpicolinat, Zinkcitrat oder Plazebo lediglich für das Picolinat eine signifikante Erhöhung der Zinkspiegel in den Haaren, im Urin und den Erythrozyten.
Anderen Untersuchungen zufolge scheint die Resorption von Zink aus Zinkgluconat mit derjenigen von Zinksulfat vergleichbar zu sein [15]. Im Rahmen einer Pharmakokinetikstudie untersuchten Neve et al. (1993) an zehn gesunden Probanden die Serum-Zinkprofile nach oraler Gabe von sechs Zubereitungsformen von Zinksulfat und Zinkgluconat, jeweils verabreicht in einer 45 mg Zink äquivalenten Dosis [36]. Untersucht wurden
- eine wässrige Lösung von Zinksulfat,
- Kapseln mit 45 mg Zink in Form von Zinksulfat ohne Hilfsstoffe,
- Kapseln mit jeweils 15 mg Zink in Form von Zinksulfat, ohne Hilfsstoffe,
- Kapseln mit 45 mg Zink in Form von Zinkgluconat, ohne Hilfsstoffe,
- Kapseln mit jeweils 15 mg Zink in Form von Zinkgluconat, mit den Hilfsstoffen Weizenstärke, Lactose, Siliciumdioxid und Magnesiumstearat,
- magensaftresistente Tabletten mit jeweils 15 mg Zink in Form von Zinkgluconat, mit verschiedenen Hilfsstoffen, darunter Sorbitol und Polyethylenglykol.
Den Ergebnissen zufolge könnte die Verteilung der gesamten Zinkdosis auf mehrere Einzeldosen die Gesamtverfügbarkeit von Zink steigern. Offensichtlichen Einfluss hatten auch die Hilfsstoffe: Sie verzögerten die Zinkaufnahme. Das schlechteste Ergebnis lieferten die magensaftresistenten Tabletten. Insgesamt stellten die Autoren eine geringfügig bessere Bioverfügbarkeit von Zinkgluconat gegenüber Zinksulfat fest.
Zinkorotat
Orotsäure ist gebräuchlich als Komplexpartner für Mineralstoffe, so zum Beispiel Magnesium, Calcium, Eisen, Kupfer, Lithium oder Zink. Die Verbindung Orotsäure selbst wurde im Rahmen der Kommission B des ehemaligen Bundesgesundheitsamtes negativ monographiert, vor allem aufgrund des unzureichenden Nachweises der klinischen Wirksamkeit [37].
Obwohl Zinkorotat seit Jahren als Mittel zur Zinksupplementation vertrieben wird, scheint die Bioverfügbarkeit vergleichsweise schlecht dokumentiert. Schölmerich et al. (1987) sehen bei dieser Verbindung keinen relevanten Vorteil gegenüber Zinksulfat [15].
Andermann und Dietz (1982) untersuchten die Bioverfügbarkeit und Pharmakokinetik von Zinkpantothenat (s. u.), Zinksulfat und Zinkorotat nach parenteraler und oraler Gabe am Kaninchen [35]. Zinkorotat zeigte nach parenteraler Gabe eine schnellere Distributions- und Eliminationsphase als die beiden anderen Substanzen, deren Unterschiede nicht signifikant waren. Dagegen wurde Zinkorotat bei oraler Gabe langsamer resorbiert als die beiden anderen Salze.
Zinkpantothenat
Guillard et al. (1978) suchten nach einem Zinksupplement, das besser verträglich ist als das zum damaligen Zeitpunkt bei Acrodermatitis üblicherweise eingesetzte Zinksulfat. Sie verglichen hierzu die Bioverfügbarkeit von Zinksulfat und Zinkpantothenat (in Deutschland nicht im Handel) nach parenteraler und oraler Gabe am Kaninchen [34]. Die Ergebnisse waren für beide Substanzen nahezu identisch. Nach oraler Applikation weist Zinkpantothenat eine geringfügig schlechtere Bioverfügbarkeit auf. Zu ähnlichen Ergebnissen kamen auch Andermann und Dietz (1982), die zusätzlich Zinkorotat (s. o.) in die Bestimmung der Bioverfügbarkeit aufnahmen [35].
Zinkaspartat
Die Verwendung von Zinkaspartat als Zinkquelle geht möglicherweise auf eine ältere tierexperimentelle Arbeit an Kaninchen zurück. Dort wurde eine bessere Resorption von Zinkaspartat im Vergleich zu Zinkchlorid und Zinksulfat festgestellt [38]. Im Gegensatz zu diesen Befunden stehen die Versuche von Schwarz und Kirchgessner (1975) an isoliertem Rattendarm. Sie fanden gegenüber der Kontrolle keine verbesserte Zinkaufnahme durch Asparaginsäure als Komplex-Ligand [39]. Dagegen erhöhte im gleichen Versuch die Zugabe des Liganden Histidin die Zinkaufnahme um den Faktor 10.
Schölmerich et al. (1987) stellten für Zinkaspartat keinen signifikanten Vorteil gegenüber Zinksulfat oder gar Zink-Histidin fest [15]. Duisterwinkel et al. (1986) kamen ein Jahr zuvor bei einer klinischen Untersuchung der Bioverfügbarkeit eines Handelspräparates mit Zinkaspartat zu einem negativen Urteil: In einer Studie an 7 gesunden Probanden fanden sie bei Gabe von Zinkaspartat in einer Dosis von 50 mg Zink-Äquivalenten keine signifikant erhöhten Plasmaspiegel, weder bei Nüchterneinnahme noch bei Einnahme zum Essen [40]. Aus ihren Ergebnissen folgerten die Autoren, dass Zink aus Zinkaspartat in magensaftresistenter Zubereitung nicht resorbiert wird. Dies bestätigte ein Kontrollversuch mit zerstoßenen Tabletten an einem Probanden: Er erbrachte eine Erhöhung der Plasma-Zinkkonzentration in der gleichen Größenordnung, wie sie auch bei Einnahme von Zinksulfat beobachtet wird.
Neuere tierexperimentelle oder klinische Studien zu Zinkaspartat scheinen nicht veröffentlicht worden zu sein, obwohl es seit Jahren als Mittel für die Zinksupplementation im Gebrauch ist.
Zink-Lysin und Zink-Methionin
Rojas et al. (1995) sowie Wedekind et al. (1994) verglichen in tierexperimentellen Arbeiten am Schaf und am Schwein die Bioverfügbarkeit von Zinkoxid und Zinksulfat mit der von Zink-Lysin und Zink-Methionin (beide nicht im Handel) [28, 29]. Dabei wurde Zink-Lysin gegenüber Zinksulfat je nach Versuch als besser oder aber deutlich schlechter bioverfügbar eingestuft.
Wedekind et al. stellten im Tierversuch an Hühnern fest, dass Zink-Methionin im Vergleich zu Zinksulfat je nach Futtergrundlage eine relative Bioverfügbarkeit von 117% bzw. 177% hat [26].
Zink-Histidin
Die Steigerung der Bioverfügbarkeit von Zink durch Komplexierung mit Histidin war Gegenstand einer Vielzahl tierexperimenteller und klinischer Untersuchungen. Sieht man von dem üblichen Referenzpräparat Zinksulfat ab, so kann man Zink-Histidin als die am besten untersuchte Zinkform betrachten. Besonders die resorptionsfördernde Wirkung von Histidin, die physiologische Co-Transportfunktion und die gute Verträglichkeit des Komplexes aus Zink und Histidin wird in der medizinischen Literatur immer wieder herausgestellt.
Die WHO weist in einer Studie zur Kinderernährung explizit auf Histidin als resorptionsfördernden Komplexpartner für Zink hin, ebenso das britische Gesundheitsministerium in der Ausarbeitung einer Expertengruppe [5, 23].
Experimentelle Arbeiten zur Resorption von Zink-Histidin
An Muscheln lässt sich der Einfluss verschiedener im Wasser gelöster Zinkformen auf die Resorption relativ einfach nachweisen. So nimmt die Miesmuschel (Mytilus edulis) Zink in Form von Zink-Histidin schneller und effektiver auf als Zink, das an andere Liganden gebunden ist [41].
Zink moduliert die Neurotransmission in Hirngebieten, die mit kognitiven Funktionen im Zusammenhang stehen: Zinkmangel schwächt das Kurzzeitgedächtnis erheblich. Dieser Effekt kann indirekt zur Bewertung der Bioverfügbarkeit herangezogen werden. So wurde in einer aktuell erschienenen tierexperimentellen Untersuchung an Ratten der Einfluss von Zinkchlorid versus Zink-Histidin auf künstlich erzeugte Zinkmangelzustände untersucht. Nach Gabe von Zink-Histidin normalisierte sich die Hirnfunktion signifikant schneller. Zink-Histidin wies eine höhere Bioverfügbarkeit auf als Zinkchlorid [42].
Wedekind et al., Van Wouwe et al. und Buxani-Rice et al. [26, 43 - 45] zeigten, dass an Aminosäuren gebundenes Zink eine bessere Bioverfügbarkeit aufweist als anorganische Zinksalze. Dies gilt tierexperimentellen Befunden zufolge insbesondere für Histidin [39, 45 - 48].
Co-Transport von Zink und Histidin
Nach Wapnir et al. werden Zink und Histidin in Form eines Co-Transportes resorbiert [16, 48]. Horn et al. (1995) untersuchten die Stereospezifität des Co-Transportes von Zink und Histidin bzw. Cystein an Erythrozyten der Ratte und des Menschen. Während die l-Formen beider Aminosäuren die Aufnahme von Zink in die Erythrozyten der Ratte signifikant steigerten, galt dies an den humanen Erythrozyten nur für l-Histidin, nicht aber für l-Cystein. Das Enantiomer, d-Histidin, hemmte die Zinkaufnahme in beiden Modellen, sowohl den tierischen als auch den humanen Erythrozyten [50].
Diese Ergebnisse sind konsistent mit den Befunden von Aiken et al. (1992), die unter Verwendung von radioaktivem 65Zn2+ an Ratten-Erythrozyten für Histidin gleichfalls eine dosisabhängige und stereospezifische Stimulation der Zinkaufnahme nachgewiesen hatten [49].
Umstritten schien lange Zeit der Effekt hoher Dosen von Histidin auf den Zinkspiegel. Bei hohen Konzentrationen von Histidin gegenüber Zink im Verhältnis 130 : 1 hemmt die Aminosäure die Resorption [49]. Van Wouwe et al. (1990) erzielten bei einem molaren Verhältnis von Histidin zu Zink im Bereich von 1750 : 1 an Mäusen eine verstärkte Ausscheidung von Zink [51], ebenso wie Wensink et al. (1988) an Ratten bei einem Verhältnis von 4000 : 1 [52] und Henkin et al. (1975) im klinischen Experiment mit einem Verhältnis von 2000 bis 3500 : 1 [24].
Van Wouwe et al. (1990) weisen darauf hin, dass diese Befunde für physiologische Verhältnisse ohne Relevanz sind: Selbst unter Extrembedingungen wird ein molares Histidin-Zink-Verhältnis von 2000 : 1 nicht erreicht werden können. Außerhalb des physiologisch relevanten Bereiches lagen auch Mengenverhältnisse zwischen 100 und 700 : 1, die in verschiedenen Studien an Ratten und Mäusen [51, 53, 54], aber auch am Menschen [24, 55] Einsatz fanden. In keiner dieser Arbeiten wurde eine Negativbeeinflussung des Zinkhaushaltes durch Histidin festgestellt.
Mit den als Zinksupplement verfügbaren Zink-Histidin-Formulierungen im Verhältnis Histidin : Zink = 2 : 1 ist eine verstärkte Ausscheidung von Zink und somit die Induktion eines Zinkmangels auf keinen Fall zu erwarten. Im Gegenteil stellten Van Wouwe et al. (1990) unter gleichzeitiger subkutaner Verabreichung von radioaktivem 65Zn2+ und Histidin an Mäusen eine stärkere Akkumulation von Zink in verschiedenen Körpergeweben fest, vor allem im Pelz der Versuchstiere und in manchen Hirnarealen [51].
Die Hypothese eines Co-Transportes wurde bereits 1960 von der Arbeitsgruppe um Sivarama Sastry aufgestellt [56]. Die von dieser Arbeitsgruppe gemessene Spezifität lag zwar um eine Zehnerpotenz niedriger als die Befunde von Aiken et al. (1992) [49]. Diese Diskrepanz erklärt sich vermutlich jedoch aus der Verwendung eines Histidin-Racemates anstelle von l-Histidin, und der damit verbundenen Hemmung der Aufnahme von Zink-Histidin in die Erythrozyten durch d-Histidin.
Einfluss von Aminosäuren auf die Zinkresorption im Darm
Wapnir et al. verglichen in ihren In-vivo-Untersuchungen am Ratten-Ileum den Einfluss niedermolekularer Liganden auf die Zinkresorption [16, 48]. Organische Säuren wie Galakturonsäure, Glucuronsäure oder Citronensäure hatten einen fördernden Einfluss auf die Zinkaufnahme. Während Picolinsäure, die für die gute Verfügbarkeit von Zink in Muttermilch verantwortlich gemacht wurde, keine besonders herausragende Förderung der Zinkresorption verursachte, hatten die Aminosäuren Glutaminsäure, Glycin und Histidin ausgeprägte Effekte.
Ziel einer pharmakologischen Untersuchung von Schwarz und Kirchgessner aus dem Jahr 1975 war die Messung des Einflusses verschiedener Aminosäuren auf die Aufnahme von Zink aus dem Rattendarm [39]. Glycin, Asparaginsäure, Tyrosin, Lysin und Methionin veränderten den intestinalen Zink-Durchtritt im Vergleich zur Kontrolle nicht, Valin, Phenylalanin und Tryptophan nur wenig. Ein leicht verbesserter Zink-Durchtritt wurde für Threonin festgestellt. Dagegen erhöhten Histidin und Cystein den intestinalen Zink-Durchtritt um den Faktor 10 bzw. 14.
Wapnir et al. [57] verglichen den Einfluss verschiedener Aminosäuren auf die Zinkresorption der Ratte. Tryptophan, Prolin und Histidin wirkten sich förderlich aus, dagegen erzielten die Autoren mit Cystein keinen signifikanten Effekt. Die Zinkresorption hängt vom pH-Wert ab. Im Dünndarm (pH 6,5 bis 7,6) scheint Histidin optimal zu wirken, dagegen ist im Colon eine Abhängigkeit der Zinkaufnahme von Histidin nicht mehr nachweisbar.
Steuerung des Zinkhaushaltes durch Zink-Histidin
Zink liegt im Serum zu einem Drittel fest an alfa2-Makroglobulin und zu ca. zwei Dritteln in loser Form an Albumin gebunden vor. Ein kleiner Prozentsatz (ca. 2%) des Zinks im Serum ist an freie Aminosäuren gebunden, unter denen vor allem Histidin und Cystein die physiologischen Transportfähren darstellen [49, 53, 58, 59].
Histidin und Cystein sind in der Lage, Zink aus der Albuminbindung zu lösen und so einer Verteilung an die Orte des Bedarfs oder der Ausscheidung überschüssiger Mengen zuzuleiten. Harraki et al. untersuchten die Zinkaufnahme in Fibroblasten anhand unterschiedlicher Mischungen von radioaktivem Zink und den Aminosäuren Taurin, Glutamin und Histidin [60]. Histidin steigert diesen Ergebnissen zufolge die Bioverfügbarkeit von Zink durch Herauslösen des Metalls aus der Plasma-Albuminbindung.
bzw. in Form von Zinksulfat. Werte nach [15].
Histidin und Cystein sind nach Giroux und Henkin (1972) die einzigen physiologisch relevanten Aminosäure-Liganden für Zink im Plasma [58]. An Aminosäuren gebundenes Zink stellt zwar nur 1 bis 3% der Gesamtmenge von Zink im Plasma (s. o.). Dennoch ist diese scheinbar geringe Menge im Sinne des Massenwirkungsgesetzes von ausschlaggebender Bedeutung für die renale Elimination von überschüssigem Zink [61]. Bestätigt durch Aiken et al. (1992) [49], fanden Gachot et al. (1991) im Rahmen von Untersuchungen an kultivierten proximalen Nierenzellen des Kaninchens zwei Eliminationsprozesse für Zink: einen unspezifischen Transport von freien Zinkionen durch ein Carrier-vermitteltes System sowie einen spezifischen Weg durch einen Natrium-abhängigen Co-Transport mit Histidin oder Cystein [61].
Dieser letztere Eliminationsweg könnte ein Schutzmechanismus des Organismus vor Hyperzinkämien sein, wie sie zum Beispiel bei katabolen Erkrankungen durch Abbau von Muskelmasse auftreten können. Die renale Ausscheidung von überschüssigem Zink wird dabei durch die gleichfalls aus dem Muskel freigesetzten Aminosäuren Cystein und Histidin gesteuert [62, 63].
Klinische Untersuchungen
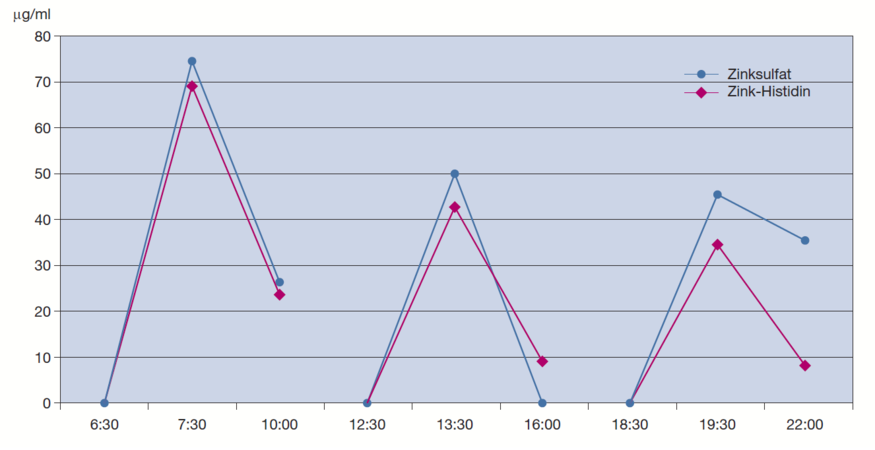
Schölmerich et al. untersuchten zunächst die Zeitabhängigkeit der Zinkaufnahme aus Zink-Histidin an Patienten mit histologisch nachgewiesener Leberzirrhose im Vergleich zu gesunden Probanden [64].* Patienten mit Leberzirrhose resorbieren dabei relevante Mengen von Zink-Histidin, wenn auch - erkrankungsbedingt - weniger als die Gesunden.
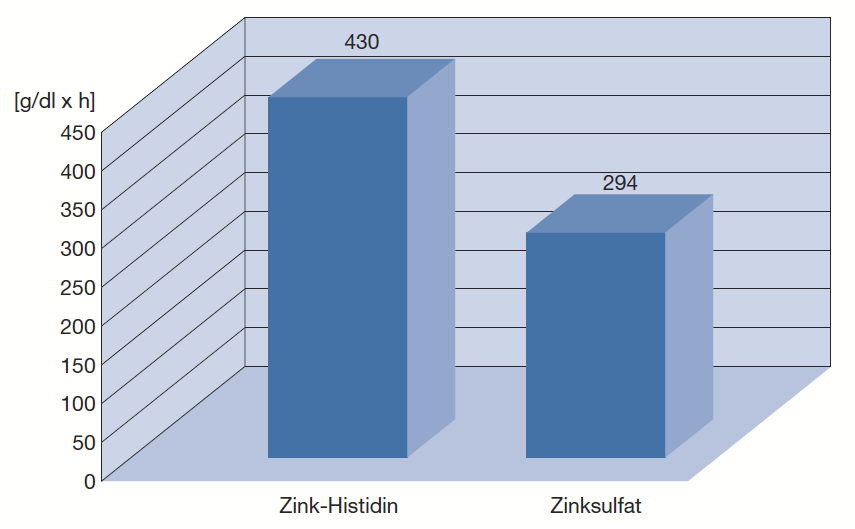
Schölmerich et al. untersuchten weiterhin im Rahmen einer Bioverfügbarkeitsstudie an gesunden Probanden die Unterschiede in der Resorption von Zink-Histidin im Vergleich zu Zinksulfat [15].* Gegenüber Zinksulfat steigerte Zink-Histidin die Aufnahme von Zink um 30 bis 40%. Die Gabe von 15 mg in Form von Zink-Histidin hatte den gleichen Effekt auf den Serum-Zinkspiegel wie die Einnahme von 45 mg Zink in Form von Zinksulfat.
Nach Gabe von Zink-Histidin beobachteten Schölmerich et al. schnelle Umverteilungsprozesse aus dem Plasma in die Gewebe hinein. Da die renale Zinkausscheidung mit 15 mg Zink-Histidin nicht gesteigert wurde, gehen sie von einer noch besseren Bioverfügbarkeit aus, als es die Werte andeuten. (Versuche mit radioaktiv markiertem Zink zur Verifizierung dieser Vermutung verbieten sich in klinischen Experimenten aufgrund der langen Halbwertszeit von 245 Tagen für Zink-Radioisotope.) Bei Einnahme von Zinksulfat und Zinkacetat in hohen Dosen wurde über Unverträglichkeiten im Gastrointestinaltrakt und Erosionen der Magenschleimhäute als Nebenwirkung der Therapie berichtet [5, 65].
Samman und Roberts (1987) [66] berichteten über das Auftreten von Übelkeit, Magenkrämpfen, Durchfall und Kopfschmerzen bei 84% gesunder weiblicher Probanden, die sechs Wochen lang täglich jeweils 150 mg Zink in Form von Zinksulfat eingenommen hatten. Bei einer Reduktion der Zinkzufuhr durch Verabreichung des besser bioverfügbaren Zink-Histidins sind diese Nebenwirkungen nicht zu erwarten.
Fleischkonsum und Zinkstatus
Das Deutsche Institut für Ernährungsmedizin und Diätetik (DIET e.V.) warnte unlängst vor einer Mangelversorgung mit essenziellen Nährstoffen, ausgelöst durch eine plötzliche Änderung der Ernährungsgewohnheiten [1]. Aufgrund der aktuellen Krisen in der Landwirtschaft (BSE, Maul-undKlauen-Seuche, Massentierhaltung, Antibiotika und andere Arzneimittel als Masthilfen) ist in der Bevölkerung ein rückläufiger Fleischkonsum zu beobachten, der sich auch auf die Versorgung mit essenziellen Nährstoffen auswirken kann. Von diesen Entwicklungen vor allem betroffen ist das Spurenelement Zink.
Der Zinkstatus bei Menschen wird in erster Linie von drei Faktoren beeinflusst [2]:
- Ausmaß der Zinkzufuhr,
- gesteigerte Zinkverluste oder höherer Zinkbedarf bei bestimmten Erkrankungen wie Diabetes, chronischen Entzündungen und dermatologischen Erkrankungen,
- gesteigerte Zufuhr von Inhibitoren der Zinkaufnahme mit der Nahrung.
Fleisch ist unsere Hauptquelle für die Zinkzufuhr. Der plötzliche Verzicht auf Fleisch geht zumeist mit einer Erhöhung der Zufuhr vegetabiler Nahrung einher und somit einer erhöhten Zufuhr von Phytaten, was die Zinkversorgung zusätzlich negativ beeinflusst.
Besonders für Risikogruppen, zu denen neben Kranken auch schwangere und stillende Frauen, Kinder in Wachstumsphasen, Senioren, Rekonvaleszente und Sportler zählen, empfiehlt sich die Substitution von Zink mit einem als Arzneimittel zugelassenen und in seiner pharmazeutischen Qualität gesicherten Zinkpräparat. Wenig bekannt ist bislang, dass sich Zinkpräparate je nach eingesetzter Zinkform teilweise erheblich in ihrer Bioverfügbarkeit unterscheiden.
Problematische Bioverfügbarkeit
Die Bioverfügbarkeit von Zink variiert beträchtlich. Insbesondere pflanzliche Kost ist zumeist eine schlechte Zinkquelle. Zink bildet mit den in Pflanzen enthaltenen Ballaststoffen, vor allem der Phytinsäure, schwerlösliche Komplexe, die einer Resorption entzogen sind. Verantwortlich für diesen Effekt sind insbesondere Phytate vom Typ des Inositolpenta- bzw. -hexaphosphates. Die Zufuhr von Zink kann demzufolge auch dann ungenügend sein, wenn Analysen der pflanzlichen Kost ausreichende Zinkgehalte andeuten [2]. Auch Oxalat oder Tannine [4] sowie Schwermetalle wie Cadmium [2] können die Zinkaufnahme behindern. Die Beeinflussung durch Cadmium dürfte in der Praxis allerdings keine relevante Rolle spielen.
In der medizinischen Literatur wird eine gegenseitige Resorptionsbeeinflussung verschiedener Mineralstoffe diskutiert. Eine postulierte Interaktion mit Calcium erschien unter verschiedenen Versuchsbedingungen eher zweifelhaft [2]. Vergleichbares gilt für die Interaktion zwischen Kupfer und Zink [2]. Trotzdem wird bei gleichzeitiger Einnahme von Zink und Kupfer vor sekundärem Kupfermangel gewarnt [5]. Die Relevanz dieser Wechselwirkung ist eher zweifelhaft [3]. So beobachteten Sullivan et al. [6] im Rahmen ihrer Bioverfügbarkeitsstudie mit Zinksulfat keinerlei Beeinflussung des Kupfer-, Calcium- und Magnesiumspiegels trotz Verabreichung der relativ hohen Dosis von 50 mg Zink in einer Einmaldosis. Dennoch empfiehlt es sich, die Einnahme von Kupfer-Supplementen zeitlich von der von Zinksupplementen zu trennen.
Hemmung der Zinkaufnahme durch Eisen?
Auch die bei gleichzeitiger Gabe von Eisen und Zink beobachtete Hemmung der Zinkresorption tritt möglicherweise nur unter den artifiziellen Bedingungen des Probandenversuchs auf. Solomons und Jacob [7] stellten fest, dass hochdosiertes anorganisches Eisen die Zinkaufnahme hemmte, gemessen an der Veränderung des Plasma-Zinkspiegels innerhalb von vier Stunden nach oraler Einnahme von Zink. Die Untersucher verabfolgten an erwachsene, nüchterne Probanden jeweils 25 mg Zink in Form von Zinksulfat und Eisen in einer Dosis von 25, 50 oder 75 mg. Bereits bei 25 mg Eisen war die Zinkaufnahme signifikant niedriger als bei den Kontrollpersonen.
Unter physiologischeren Bedingungen scheint dieser Effekt jedoch weniger stark ausgeprägt zu sein [8] und bei Zugabe von Histidin zur Testdosis sogar dann aufgehoben, wenn das Dosisverhältnis zwischen Eisen und Zink mit 25 : 1 in unphysiologischen Bereichen lag. Dieses Ergebnis wurde mittlerweile in verschiedenen tierexperimentellen und klinischen Untersuchungen mit Radioisotopen bestätigt (z. B. [9, 10]).
Eisen und Zink können in pharmazeutischen Zubereitungen bedenkenlos gemeinsam eingesetzt werden [2]. Auch die Langzeiteinnahme von Eisen- und/oder Zinksupplementen hat keinen negativen Einfluss auf den Versorgungsstatus beider Mineralstoffe [2, 8].
Bessere Zinkresorption durch Kopplung an Aminosäuren
Gute Zinkquellen sind zumeist tierische Produkte, zum Beispiel Rindfleisch oder Milchprodukte. Die Proteine in der Nahrung beeinflussen die Zinkaufnahme, wobei verschiedene Proteine durchaus unterschiedliche Effekte zur Folge haben können. So ist zum Beispiel für Casein eine geringgradige Hemmung der Zinkaufnahme bekannt.
Dagegen zeigen bestimmte Aminosäuren, insbesondere Methionin, Cystein oder Histidin, einen deutlich fördernden Einfluss auf die Zinkaufnahme [2]. Der positive Einfluss von Nahrungsproteinen tierischen Ursprungs wird denn auch weniger auf den Einfluss des Eiweißmoleküls selbst als vielmehr auf die daraus freigesetzten freien Aminosäuren zurückgeführt. Diese Aminosäuren können Zink auch in Anwesenheit von Phytaten in gelöster, resorbierbarer Form halten - ein Effekt, der auch für Oligopeptide aus Casein bereits nachgewiesen wurde.
Muttermilch als Zinkquelle
Hinsichtlich der Bioverfügbarkeit von Zink bestehen zwischen verschiedenen Nahrungsmitteln erhebliche Unterschiede. Da Kinder in Wachstumsphasen sensibel auf die Zinkversorgung reagieren, ist die Beachtung des Zinkstatus in der Kinderernährung besonders wichtig. Für Säuglinge ist die Muttermilch die beste Zinkquelle - vorausgesetzt, die Mutter weist einen ausreichenden Zinkstatus auf. So wurde anhand radioaktiv markierter Zinkzusätze in tierexperimentellen Studien an Ratten festgestellt, dass die Zink-Bioverfügbarkeit aus menschlicher Muttermilch bei 28% liegt, aus Kuhmilch bei 15% und aus Soja-Formula-Nahrung bei nur 10% [11].
Evans et al. machten eine Komplexbildung mit Picolinsäure für die gute Bioverfügbarkeit von Zink aus Muttermilch verantwortlich [12]. Während Picolinsäure, die mit Zink einen Chelatkomplex bilden kann, in der Muttermilch in einer Konzentration von 308 Mikromol/l vorkommt, beträgt die Konzentration in Kuhmilch lediglich 20 Mikromol/l oder weniger. Picolinsäure verbessert die Bioverfügbarkeit von Zink: Barrie et al. [13] fanden im Rahmen eines vierwöchigen Probandenversuchs eine Erhöhung des Zinkspiegels in den Haaren, im Urin und den Erythrozyten.
Allerdings reicht einigen Autoren zufolge die Konzentration von Picolinsäure in der Muttermilch nicht aus, um die Verfügbarkeit von Zink aus dieser Quelle befriedigend zu erklären. So weist Zinkpicolinat gegenüber dem Komplex Zink-Histidin eine deutlich unterlegene Verfügbarkeit auf [14-16]. Histidin als essenzielle Aminosäure ist gleichfalls Bestandteil der Muttermilch und könnte einen zusätzlichen Beitrag für die gute Bioverfügbarkeit von Zink in Muttermilch leisten [11].
Resorptionsverbesserer für Zink
Verschiedene niedermolekulare Substanzen wurden auf ihre resorptionsfördernde Wirkung für Zink untersucht:
- Ascorbinsäure, deren positiver Einfluss auf die Eisenaufnahme gut bekannt ist, bewirkte in Dosen zwischen 0,5 und 2 Gramm nach Gabe von 110 mg Zinksulfat-Heptahydrat, entsprechend 25 mg Zink, keine Veränderung der Zinkaufnahme [17].
- Auch für Citronensäure waren die Ergebnisse unbefriedigend. Der Zusatz von Citrat zu Lebensmitteln kann zwar unter Umständen die Resorption von Zink verbessern [18]. Für eine kontrollierbare Supplementation mit Zinkpräparaten kommt Zinkcitrat jedoch nicht infrage. Barrie et al. [13] konnten zudem bei Gabe von Zinkcitrat keine Veränderung des Serum-Zinkspiegels nachweisen.
- EDTA (Ethylendiamintetraacetat), eine Chelat-bildende Substanz, die zur Komplexierung von Metallionen eingesetzt wird, wurde bereits frühzeitig als potenzielle Zinkfähre in Tierversuchen an Hühnern getestet [19, 20]. Allerdings stellte sich in der Folge heraus, dass komplexiertes Zink-EDTA zwar gut aus dem Darmlumen in die Enterozyten aufgenommen wird, von hier aus aber nicht die basolaterale Membran durchdringen kann [21]. Um für den Organismus verwertbar zu sein, müssen die Zinkionen daher zunächst umkomplexiert werden.
- Aminosäuren sind gute Zink-Chelatoren, insbesondere Histidin [2, 5, 22, 23]. Die Eignung von Zink-Histidin als Zinklieferant wurde in klinischen Studien bewiesen (s. u.). Dabei kommt es auf das Mengenverhältnis zwischen Zink und Histidin an: Extrem hohe Mengen an Histidin (60 Gramm pro Tag), die weder in der Nahrung noch in Zinksupplementen vorkommen, können sogar zu einer verstärkten Ausscheidung von Zink über den Urin führen [24]. Zink-Histidin wird jedoch bei hoher Dosierung renal ausgeschieden [25], was eine - beabsichtigte oder versehentliche - Überdosierung ausschließt.
Zusammenfassung
Zink zählt zu den essenziellen Nährstoffen mit problematischer Bioverfügbarkeit. Aufgrund der modernen Ernährungsgewohnheiten sind grenzwertige Zinkmangelzustände auch in Deutschland ein ernst zu nehmendes Problem. Risikogruppen sind insbesondere Diabetiker, Allergiker, Neurodermitiker, Psoriasiskranke, Patienten mit chronisch-entzündlichen Erkrankungen, Rekonvaleszente, Sportler, stillende und schwangere Frauen, Senioren sowie Kinder in Wachstumsphasen.
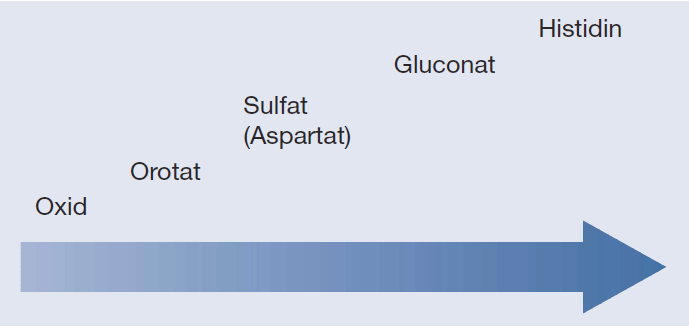
Auch wenn die unterschiedlichen Messmethoden eine abschließende Aussage nicht gestatten, kann aus der Literatur hinsichtlich der Bioverfügbarkeit von Zinkpräparaten folgende Reihenfolge abgeleitet werden: Zinkoxid << Zinkorotat < Zinksulfat < Zinkgluconat << Zink-Histidin. Zinkaspartat lässt sich in diese Reihe aufgrund mangelnder Daten nicht einsortieren.
Insbesondere Diabetiker, die krankheitsbedingt erhöhte Zinkverluste aufweisen, profitieren von einer geregelten Zinkversorgung. Zink kann bei diesen Patienten zu einem besseren Ansprechen auf Antidiabetika (verbesserte Glucosetoleranz, verminderte Insulinresistenz) sowie einem schnelleren Abheilen von Haut- und Schleimhautdefekten führen (z. B. [67-69]).
Bei Arzneistoffen mit problematischer Bioverfügbarkeit, wie es beim Zink der Fall ist, hat die Wahl eines gut bioverfügbaren Präparates einen erheblichen Einfluss auf das Erreichen des optimalen Therapiezieles.
Kastentext: Problematische Resorption
Die Zufuhr der empfohlenen Tagesdosis von 15 mg Zink kann dann unzureichend sein, wenn aufgrund der schlechten Bioverfügbarkeit des Mineralstoffs aus der jeweiligen Zinkquelle nur Bruchteile dieser Zinkdosis zur Resorption kommen [3].
Fußnote
*Prüfpräparat Curazink. Eine Kapsel enthält 94 mg Zink-Histidin bzw. 15 mg Zink.
Literatur [1] Problem Zinkmangel durch BSE-Krise in Deutschland. Gourmed 2001; 17 (1): 58 - 60. [2] Lönnerdal B. Dietary factors influencing zinc absorption. J Nutr 2000; 130 (Suppl.): 1378S - 1383S. [3] Solomons NW. Biological availability of zinc in humans. Am J Clin Nutr 1982; 35 (5): 1048 - 1075. [4] Cousins RJ. Zinc. In: Ziegler EE, Filer LJ Jr, editors. Present Knowledge in Nutrition. Washington, DC: ILSI-Press, 1996: 293 - 306. [5] Expert Group on Vitamins and Minerals. Review of Zinc. UK Department of Health Covering Note to EVM/99/18/P 1999;1 - 45. [6] Sullivan JF, Jetton MM, Burch RE. A zinc tolerance test. J Lab Clin Med 1979; 93: 485 - 492. [7] Solomons NW, Jacob RA. Studies on the bioavailability of zinc in humans: effects of heme and nonheme iron on the absorption of zinc. Am J Clin Nutr 1981; 34 (4): 475 - 482. [8] Sandström B, Davidsson L, Cederblad A, Lönnerdal B. Oral iron, dietary ligands and zinc absorption. J Nutr 1985; 115 (3): 411 - 414. [9] Davidsson L, Almgren A, Sandström B, Hurrell RF. Zinc absorption in adult humans: the effect of iron fortification. Br J Nutr 1995; 74: 417 - 425. [10] Fairweather-Tait SJ, Wharf SG, Fox TE. Zinc absorption in infants fed iron-fortified weaning food. Am J Clin Nutr 1995; 62: 785 - 789. [11] Sandström B, Keen CL, Lönnerdal B. An experimental model for studies of zinc bioavailability from milk and infant formulas using extrinsic labeling. Am J Clin Nutr 1983; 38 (3): 420 - 428. [12] Evans GW, Johnson PE. Characterization and quantitation of a zinc-binding ligand in human milk. Pediatr Res 1980; 14 (7): 876 - 880. [13] Barrie SA, Wright JV, Pizzorno JE, Kutter E, Barron PC. Comparative absorption of zinc picolinate, zinc citrate and zinc gluconate in humans. Agents Actions 1987; 21 (1 - 2): 223 - 228. [14] Johnson WT, Evans GW. Tissue uptake of zinc in rats following the administration of zinc dipicolinate or zinc histidinate. J Nutr 1982; 112 (5): 914 - 919. [15] Schölmerich J, Freudemann A, Köttgen E, Wietholtz H, Steiert B, Löhle E et al. Bioavailability of zinc from zinc-histidine complexes. I. Comparison with zinc sulfate in healthy men. Am J Clin Nutr 1987; 45 (6): 1480 - 1486. [16] Wapnir RA, Garcia-Aranda JA, Mevorach DE, Lifshitz F. Differential absorption of zinc and low-molecular-weight ligands in the rat gut in protein-energy malnutrition. J Nutr 1985; 115 (7): 900 - 908. [17] Solomons NW, Jacob RA, Pineda O, Viteri FE. Studies on the bioavailability of zinc in man. III. Effects of ascorbic acid on zinc absorption. Am J Clin Nutr 1979; 32 (12): 2495 - 2499. [18] Pabon M, Lönnerdal B. Effect of citrate on zinc bioavailability from milk, milk fractions and infant formulas. Nutr Res 1993; 13: 103 - 111. [19] Kratzer FH, Allred JB, Davis PN, Marshall BJ, Vohra P. The effect of autoclaving soybean protein and the addition of ethylenediaminetetraacetic acid on the biological availability of dietary zinc for turkey poults. J Nutr 1959; 68: 313 - 322. [20] O'Dell BL. Effect of dietary components upon zinc bioavailability. Am J Clin Nutr 1969; 22 (10): 1315 - 1322. [21] Hempe JM, Cousins RJ. Effect of EDTA and zinc-methionine complex on zinc absorption by rat intestine. J Nutr 1989; 119: 1179 - 1187. [22] Coyle P, Philcox JC, Rofe AM. Zinc in man. Clin Biochemist Rev 1998; 19: 107 - 117. [23] Fleischer Michaelsen K, Weaver L, Branca F, Robertson A. Minerals other than iron. Feeding and nutrition of infants and young children. Guidelines for the WHO European Region, with emphasis on the former Soviet countries. Kopenhagen: WHO Regional Publications, European Series, No. 87, 2000: 85 - 100. [24] Henkin RI, Patten BM, Re PK, Bronzert DA. A syndrome of acute zinc loss: cerebellar dysfunction, mental changes, anorexia, and taste and smell dysfunction. Arch Neurol 1975; 32: 745 - 751. [25] Zlotkin SH. Nutrient interactions with total parenteral nutrition: effect of histidine and cysteine intake on urinary zinc excretion. J Pediatr 1989; 114 (5): 859 - 864. [26] Wedekind KJ, Hortin AE, Baker DH. Methodology for assessing zinc bioavailability: efficacy estimates for zinc-methionine, zinc sulfate, and zinc oxide. J Anim Sci 1992; 70 (1): 178 - 187. [27] Wedekind KJ, Baker DH. Zinc bioavailability in feed-grade sources of zinc. J Anim Sci 1990; 68 (3): 684 - 689. [28] Rojas LX, McDowell LR, Cousins RJ, Martin FG, Wilkinson NS, Johnson AB et al. Relative bioavailability of two organic and two inorganic zinc sources fed to sheep. J Anim Sci 1995; 73 (4): 1202 - 1207. [29] Wedekind KJ, Lewis AJ, Giesemann MA, Miller PS. Bioavailability of zinc from inorganic and organic sources for pigs fed corn-soybean meal diets. J Anim Sci 1994; 72 (10): 2681 - 2689. [30] Henderson LM, Brewer GJ, Dressman JB, Swidan SZ, DuRoss DJ, Adair CH et al. Effect of intragastric pH on the absorption of oral zinc acetate and zinc oxide in young healthy volunteers. J Parenter Enteral Nutr 1995; 19 (5): 393 - 397. [31] Hortin AE, Oduho G, Han Y, Bechtel PJ, Baker DH. Bioavailability of zinc in ground beef. J Anim Sci 1993; 71 (1): 119 - 123. [32] Keyzer JJ, Oosting E, Wolthers BG, Muskiet FAJ. Zinc absorption after oral administration of zinc sulfate. Pharm Weekbl [Sci Ed] 1983; 5: 252 - 253. [33] Pohit J, Saha KC, Pal B. A zinc tolerance test. Clin Chim Acta 1981; 114: 279 - 281. [34] Guillard O, Saux MC, Hazane C, Dumas C, Courtois P. Pharmacocinétique comparee du sulfate et du pantothénate de zinc. Ann Pharm franć 1978; 36 (11 - 12): 669-676. [35] Andermann G, Dietz M. The bioavailability and pharmacokinetics of three zinc salts: zinc pantothenate, zinc sulfate and zinc orotate. Eur J Drug Metab Pharmacokinet 1982; 7 (3): 233 - 239. [36] Neve J, Hanocq M, Peretz A, Abi Khalil F, Pelen F. Etude de quelques facteurs influencant la biodisponibilité du zinc dans les formes pharmaceutiques à usage oral. J Pharm Belg 1993; 48 (1): 5 - 11. [37] Monographie: Orotsäure. Bundesanz vom 10. Juni 1989; 106. [38] Kruse-Jarres JD, Herr D, Vocke G, Baur B, Baur J, Waldmann D. Die Resorption von Zink und Magnesium durch den Intestinaltrakt. In: Henschel WF, Editor. Die Rolle von Kalium-Magnesium-Aspartat in der operativen Medizin und Intensivtherapie: 10 Jahre Erfahrungen mit Inzolen. Stuttgart: Schattauer-Verlag, 1977: 49 - 59. [39] Schwarz FJ, Kirchgessner M. Zum Einfluß organischer Liganden auf die Zn-Absorption in vitro. Z Tierphysiol Tierernährg Futtermittelkde 1975; 35: 257 - 266. [40] Duisterwinkel FJ, Wolthers BG, Koopman BJ, Muskiet FAJ, Van der Slik W. Bioavailability of orally administered zinc, using Taurizine. Pharm Weekbl [Sci Ed] 1986; 8: 85 - 88. [41] Vercauteren K, Blust R. Bioavailability of dissolved zinc to the common mussel Mytilus edulis in complexing environments. MEPS 1996; 137: 123 - 132. [42] Keller KA, Chu Y, Grider A, Coffield JA. Supplementation with L-histidine during dietary zinc repletion improves short-term memory in zinc-restricted young adult male rats. J Nutr 2000; 130: 1633 - 1640. [43] Buxani-Rice S, Ueda F, Bradbury MW. Transport of zinc-65 at the blood-brain barrier during short cerebrovascular perfusion in the rat: its enhancement by histidine. J Neurochem 1994; 62 (2): 665 - 672. [44] Van Wouwe JP, Hoogenkamp S, Van den Hamer CJ. Histidine supplement and Zn status in Swiss random mice. Biol Trace Elem Res 1989; 22 (1): 35 - 43. [45] Van Wouwe JP, Veldhuizen M. Growth characteristics in laboratory animals fed zinc-deficient, copper-deficient, or histidine-supplemented diets. Biol Trace Elem Res 1996; 55 (1 - 2): 71 - 77. [46] Schwarz FJ, Kirchgessner M. Tierexperimentelle Untersuchungen zur Zn-Absorption bei verschiedenen Dünndarmabschnitten und Zn-Verbindungen. Nutr Metabol 1975; 18 (3): 157 - 166. [47] Suso FA, Edwards HMJ. Binding of EDTA, histidine and acetylsalicylic acid to zinc-protein complex in intestinal content, intestinal mucosa and blood plasma. Nature 1972; 236: 230 - 232. [48] Wapnir RA, Khani DE, Bayne MA, Lifshitz F. Absorption of zinc by the rat ileum: effects of histidine and other low-molecular-weight ligands. J Nutr 1983; 113 (7): 1346 - 1354. [49] Aiken SP, Horn NM, Saunders NR. Effects of amino acids on zinc transport in rat erythrocytes. J Physiol 1992; 445: 69 - 80. [50] Horn NM, Thomas AL, Tompkins JD. The effect of histidine and cysteine on zinc influx into rat and human erythrocytes. J Physiol 1995; 489.1: 73 - 80. [51] Van Wouwe JP, Hoogenkamp S, Van den Hamer CJ. A histidine supplement and regulation of the zinc status in Swiss random mice. Biol Trace Elem Res 1990; 24 (3): 207 - 216. [52] Wensink J, Van den Hamer CJ. Effect of excess dietary histidine on rate of turnover of 65Zn in brain of rat. Biol Trace Elem Res 1988; 16 (2): 137 - 150. [53] Freeman RM, Taylor PR. Influence of histidine administration on zinc metabolism in the rat. Am J Clin Nutr 1977; 30 (4): 523 - 527. [54] Harvey PW, Hunsaker HA, Allen KG. Dietary L-histidine-induced hypercholesterolemia and hypocupremia in the rat. J Nutr 1981; 111 (4): 639 - 647. [55] Schechter PJ, Prakash NJ. Failure of oral L-histidine to influence appetite or affect zinc metabolism in man: a double-blind study. Am J Clin Nutr 1979; 32 (5): 1011 - 1014. [56] Sivarama Sastry K, Visvanathan L, Ramalah A, Sarma PS. Studies on the binding of 65Zn by equine erythrocytes in vitro. Biochem J 1960; 74: 561 - 567. [57] Wapnir RA, Stiel L. Zinc intestinal absorption in rats: specificity of amino acids as ligands. J Nutr 1986; 116 (11): 2171 - 2179. [58] Giroux EL, Henkin RI. Competition for zinc among serum albumin and amino acids. Biochim Biophys Acta 1972; 273 (1): 64 - 72. [59] Prasad AS, Oberleas D. Binding of zinc to amino acids and serum proteins in vitro. J Lab Clin Med 1970; 76 (3): 416 - 425. [60] Harraki B, Guiraud P, Rochat M-H, Faure H, Richard M-J, Fussellier M et al. Effect of taurine, L-glutamine and L-histidine addition in an amino acid glucose solution on the cellular bioavailability of zinc. Biometals 1994; 7 (3): 237 - 243. [61] Gachot B, Tauc M, Morat L, Poujeol P. Zinc uptake by proximal cells isolated from rabbit kidney: effects of cysteine and histidine. Pflügers Arch 1991; 419 (6): 583 - 587. [62] Abu-Hamdan DK, Migdal SD, Whitehouse R, Rabbani P, Prasad AS, McDonald FD. Renal handling of zinc: effect of cystein infusion. Am J Physiol 1981; 241: F487 - F494. [63] Yunice AA, King RWJ, Kraikitpanitch S, Haygood CC, Lindeman RD. Urinary zinc excretion following infusions of zinc sulfate, cysteine, histidine, or glycine. Am J Physiol 1978; 235 (1): F40 - F45. [64] Schölmerich J, Krauss E, Wietholtz H, Köttgen E, Löhle E, Gerok W. Bioavailability of zinc from zinc-histidine complexes. II. Studies on patients with liver cirrhosis and the influence of the time of application. Am J Clin Nutr 1987; 45 (6): 1487 - 1491. [65] Henderson LM, Brewer GJ, Dressman JB, Swidan SZ, DuRoss DJ, Adair CH et al. Use of zinc tolerance test and 24-hour urinary zinc content to assess oral zinc absorption. J Am Coll Nutr 1996; 15 (1): 79 - 83. [66] Samman S, Roberts DC. The effect of zinc supplements on plasma zinc and copper levels and the reported symptoms in healthy volunteers. Med J Aust 1987; 146 (5): 246 - 249. [67] Song MK, Rosenthal MJ, Naliboff BD, Phanumas L, Kang KW, Tissue E. Effects of bovine prostate powder on zinc, glucose, and insulin metabolism in old patients with non-insulin-dependent diabetes mellitus. Metabolism 1998; 47 (1): 39 - 43. [68] Becker B. Metabolisches Syndrom und seine Folgeerkrankungen. Zink in der Prävention des diabetisches Fußes. Naturheilpraxis 1999; (10): 1566 - 1569. [69] Beyer D. Stellenwert des essenziellen Spurenelementes Zink in der Diabetestherapie. Gesundes Leben 2001; 78 (3): 26 - 29.
Zinkmangel ist in unserer Bevölkerung weit verbreitet. Betroffen sind u. a. Diabetiker, Allergiker, Patienten mit chronisch-entzündlichen Darmerkrankungen, Rekonvaleszente, Sportler, stillende und schwangere Frauen, Senioren sowie Kinder in Wachstumsphasen. Ernährungswissenschaftler und Mediziner empfehlen für solche Risikogruppen die Supplementation von Zink. Die einzelnen Zinkpräparate unterscheiden sich aufgrund der unterschiedlichen chemischen Zinkverbindungen allerdings erheblich in ihrer Bioverfügbarkeit.



0 Kommentare
Das Kommentieren ist aktuell nicht möglich.